К.т.н.
Мудрик В.А.
Институт
фундаментальных проблем биологии РАН, г. Пущино Московской области, Россия
ВЛИЯНИЕ МИКРОУДОБРЕНИЙ
НА ФОТОСИНТЕТИЧЕСКУЮ АКТИВНОСТЬ ЯРОВОЙ
ПШЕНИЦЫ В УСЛОВИЯХ ДЕЙСТВИЯ СТРЕСС-ФАКТОРОВ
В вегетационных опытах в 2008 [1] году изучали
влияние предпосевной обработки семян комплексом микроэлементов «Микромак» и некорневой обработки комплексом «Микроэл»
растений яровой пшеницы Курская 2038. Установлено, что в результате совместного
применения комплексов «Микромак» и «Микроэл»
на ранних стадиях развития яровой пшеницы повышается биомасса растений,
содержание хлорофилла в листьях, скорость азотфиксации и эффективность
использования растениями солнечной энергии. Наибольшая фотосинтетическая
активность растений была в фазе кущения. Максимальная скорость ассоциативной
азотфиксации растений наблюдалась в фазах выхода в трубку и колошения. Прирост
биомассы растений в фазе кущения был за счет обработки семян «Микромак». Более
значительный прирост биомассы в фазе выхода в трубку наблюдали за счет
некорневых обработок «Микроэл». Эффект действия «Микроэл» в фазе кущения
проявился в росте содержания хлорофилла в листьях растений и увеличении
скорости фотосинтеза PN,
квантового выхода фотосистемы II (ФСII) и скорости
электронного транспорта ФСII. Это в итоге
привело к повышению эффективности использования растениями энергии в диапазоне
фотосинтетически активной радиации (ФАР). Можно полагать, что увеличение
квантового выхода и скорости электронного транспорта ФСII повышает устойчивость растений к стрессам.
Фотосинтез
– процесс, обеспечивающий клетку энергией, поэтому сохранение фотосинтетической
активности в условиях физиологического стресса во многом определяет
устойчивость растения к неблагоприятным факторам окружающей среды.
В последнее время в современных методах исследования
широко используется метод индукции флуоресценции хлорофилла (ИФХ) [2]. Метод
ИФХ обладает следующими преимуществами: информативность, экспрессность,
сохранение степени нативности образца после измерений, высокая чувствительность.
В основе метода ИФХ лежат следующие принципы:
1.
Существуют три основных
пути реализации квантов света поглощенных молекулами хлорофилла –
фотохимические реакции, тепловая диссипация и флуоресценция.
2.
Все три процесса
являются конкурентными, вследствие чего изменение одного из них ведет к
противоположному направленному изменению двух других. Поэтому интенсивность
флуоресценции чувствительна к изменениям фотохимических процессов и тепловой
диссипации.
Цель
работы: определить влияние микроудобрений
«Микромак» и «Микроэл» на фотосинтетическую активность растений яровой пшеницы
в условиях стресс-факторов. Определить влияние вариантов обработки удобрениями
на фотосинтетическую активность растений яровой пшеницы в условиях изменения
влажности почвы (засухоустойчивость),
действия низких температур воздуха (холодоустойчивость), обработки растений
гербицидами и действия высоких значений солнечного излучения
(фотоингибирование).
МЕТОДИКА ИССЛЕДОВАНИЯ
Исследования проводили в условиях вегетационного
опыта с почвой с яровой пшеницей сорта
Курская 2038. Растения выращивали в сосудах, вмещающих 3 кг сухой почвы, по 5
растений на сосуд (из расчета 500 семян на 1 м2 почвы). Для
исследований использовали серую лесную почву, которая имела следующую
характеристику: содержание гумуса – 2,6%, pHKCL -
5,8; Hг и S соответственно 1,8 и 18,0 мг-экв/100 г почвы, V – 92%, подвижные
P2O5 и К2О
соответственно 6,6 и 15,6; гидролизуемый азот – 5,3 мг/100 г почвы. Опыт проводили в оранжерее при естественных
условиях фотопериода, температуры и освещенности. Семена растений обрабатывали
комплексом микроэлементов «Микромак», состоящего из двух частей: «Микромак А»
(медь, цинк, магний, никель, литий, кобальт, железо, марганец) и «Микромак Б»
(калий, хром, молибден, бор, ванадий, селен, общий фосфор).
Для некорневой подкормки использовали комплекс микроэлементов «Микроэл».
Исследовали 3 варианта выращивания растений: вариант 1 -контрольные растения без применения микроудобрений,
вариант 2 - с обработкой семян «Микромак»,
вариант 3 - обработки семян «Микромак» и внекорневая обработка «Микроэл» (дважды в течение опыта – в фазах
кущения и выхода в трубку).
В
каждом варианте было по 20 сосудов с растениями.
В течение ранних стадий вегетации (фазы
кущения и выхода в трубку) измеряли скорости фотосинтеза в расчете на единицу
площади листьев (PN) , квантовый выход и
скорости электронного транспорта фотосистемы II (ФСII) интактных
листьев. Интенсивность солнечной радиации измеряли квантометром (Quantum Meter LI-250 Light Meter, USA) в величинах
ФАР (Вт/м2).
Скорость
фотосинтеза PN в условиях оранжереи
и в контролируемых условиях климата – климатокамере WTK-1250, измеряли портативным газоанализатором с камерой
прищепкой типа LCpro (ADC BioScientific, England).
Флуоресценцию хлорофилла «а» ФСII интактных
листьев измеряли Portable Chlorophyll Fluorometer MINI-PAM (Walz, Effeltrich, Germany) [2].
Фотосистема
II (ФСII) играет
центральную роль в генерировании и регуляции электронного транспорта в
хлоропластах. Показатели, отражающие эффективность работы ФСII, являются важным компонентом мониторинга активности
фотосинтетического аппарата листьев растений. Одной из наиболее основных
характеристик комплексов ФСII является
квантовый выход фотохимического превращения энергии, осуществляемого ФСII (φФСII). Квантовый выход фотохимии ФСII определяется как соотношение количества квантов,
используемых на фотохимию к общему количеству поглощенных квантов. Различают
максимальный квантовый выход (Fv/ Fm), (где Fv =Fm –
F0) определяется следующим образом: включают
источник измеряющего света,
интенсивность которого настолько мала, что реакционные центры ФСII остаются при
этом «открытыми» и флуоресценция достигает минимального значения F0. Затем
применяют короткую вспышку света высокой интенсивности, и флуоресценция
достигает максимального значения Fm. По
разнице Fm –
F0
оценивают потенциальную эффективность фотохимии ФСII в
адаптированном к темноте состоянием. Эффективный квантовый выход (φФСII) определяли на свету при действии актиничного
(поддерживающего фотосинтез) света. Снижение уровня сигнала (тушение
флуоресценции) вызвано фотохимическим тушением флуоресценции и увеличением
тепловой диссипации в светособирающей антенне ФСII (нефотохимическое тушение флуоресценции).
Скорость
электронного транспорта ФСII (ETR) определяется как:
ETR = φФСII × ФАР ×0,5 × 0,84;
где
коэффициент 0,5 учитывает эквивалентное
распределение световой энергии между ФСII и фотосистемой I, величина коэффициента поглощения листа, принята 0,84].
Индекс Лихтенталлера [3] «Vitality index»,
т.е. индекс жизнеспособности имеет тесную корреляцию со потенциальной скоростью
фотосинтеза PN и определяется
отношением величины снижения флуоресценции с текущим значением флуоресценции. Vitality index (IV) измеряли при насыщающей интенсивности освещения
(около 500 Вт/м2 ФАР).
Влияние гербицидов на фотосинтетическую
активность растений яровой пшеницы исследовали с применением комплексного
препарата Триатлон, в состав которого входит трибенурон-метил.
Этот новый российский
гербицид предназначен для защиты зерновых культур от широкого спектра
однолетних и многолетних двудольных сорняков. Особенностью этого препарата
является высокая селективность по отношению к культуре и высокая эффективность
при низких положительных температурах (от +50 С), за счёт этого
более широкий интервал применения (от 2—3 настоящих листьев у культуры до
выхода флагового листа) по сравнению с гормональными препаратами. А благодаря
быстрому разложению в почве не представляет опасности для последующих культур,
поэтому не имеет ограничений в севообороте.
В состав Триатлона
входят три действующих вещества: 2-этилгексиловые эфиры 2,4-Д кислоты и дикамбы
кислоты, плюс трибенурон-метил. Это позволило расширить спектр действия и
усилить гербицидный эффект против наиболее распространённых сорных растений в
посевах зерновых культур, а также свести к минимуму риск возникновения
резистентности у сорняков. Кроме того, за счет присутствия в составе Триатлона
поверхностно-активных веществ обеспечивается отличная смачиваемость листовой
поверхности, а высокая способность эмульсии проникать через кутикулу даже
опушенного или покрытого восковым налетом листа позволяет молекулам
трибенурон-метила, эфира дикамбы и 2,4-Д кислоты быстро поступать и
передвигаться по сосудистой системе сорняков. Следовательно, сокращается до
одного часа опасный интервал от обработки до возможного выпадения осадков, а
проявление первых симптомов наблюдается уже через 12—18 часов после обработки.
Таких результатов нельзя достичь, если применять препараты на основе трибенурон-метила
«в чистом виде». Норма применения Триатлона - 0,5 л/га, рекомендуется применять
в фазе кущения.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Засухоустойчивость растений яровой
пшеницы
Влагообеспеченность растений корректно
оценивать по величине доступной для растений влаги в почве, в величинах
давления почвенной влаги или водного потенциала почвы – в килопаскалях (кПа) и
мегапаскалях (МПа). Такая оценка имеет преимущество перед остальными
традиционными методами, потому что оптимальная (по фотосинтезу) величина
водного потенциала почвы для данного вида растения не зависит от типа почвы, на
которых выращивают данную культуру. К сожалению, традиционный датчик измерения
давления почвенной влаги – тензиометр – имеет диапазон почвенной влаги от 0 до
-100 кПа. Поэтому, имеющимся датчиком
давления почвенной влаги – тензиометром (фирмы “Irrometer”, USA) измерения
проводили только в диапазоне влаги, в
котором находится оптимальная (по фотосинтезу) величина водного потенциала
почвы (начало кривой зависимости PN от
влажности почвы).
Влажность
почвы определяется содержанием воды в почве. Она выражается в абсолютных
величинах – измеряется в миллиметрах слоя воды
и
относительных величинах – отношение массы воды, содержащейся в
почве, к массе сухой почвы и выражается
в процентах.
Ws =Ws1 –Ws2/ Ws2 , где
Ws1 – почва
до просушивания;
Ws2 – масса
почвы после просушивания в сушильном шкафу
Выделяют пять основных
почвенно-гидрологических констант, которые широко применяются в агрономической
и мелиоративной практике:
максимальная
гигроскопическая влажность(МГ);
влажность
завядания (ВЗ);
влажность
разрыва капилляров (ВРК);
наименьшая
влагоемкость (НВ);
полная
влагоемкость (ПВ).
Максимальная
гигроскопическая влажность.
Под НВ понимается наибольшее количество
капиллярно - подвешенной влаги, которое почва способна удержать после ее
обильного увлажнения и свободного стекания избытка влаги.
Влажность
завядания (ВЗ) - влажность, при которой растения начинают обнаруживать признаки
завядания, не исчезающие при перемещении в атмосферу, насыщенную водными
парами. Это нижний предел доступной для растений влаги.
В нашей работе мы определяли содержание
воды в почве в относительных величинах
– в % абсолютно сухой почвы.
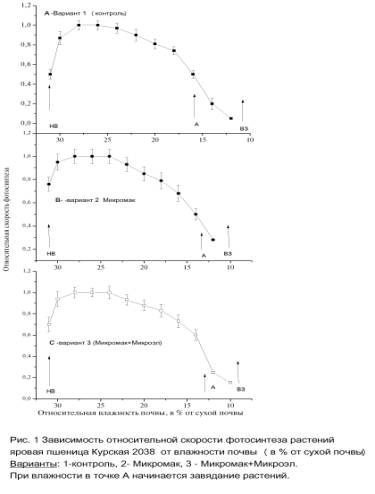
Для
всех вариантов исследованы зависимости скорости фотосинтеза от влажности почвы,
основная часть из них приведена на Рис.
1 и таблице 1.
Таблица
1. Зависимость скорости фотосинтеза PN растений яровая пшеница
Курская 2038 от влажности почвы Ws (в %
сухой почвы).
|
Влажность почвы, % |
PN, мг СО2/ дм 2 час |
|
|
Контроль |
Микромак
+ Микроэл |
|
|
32 |
- |
- |
|
31,7-
30,7 |
8,7±0,6 |
27,2±1,9 31,4±2,2 32,0±2,5 31,3±2,3 32,4±2,6 29,2±2,3 28,8±2,3 25,2±1,9 24,6±2,1 20,3±1,7 7,6±1,0 3,5±0,5 - |
|
30,3
– 29,5 |
15,1±1,1 |
|
|
28,6-
27,8 |
17,4±1,5 |
|
|
25,7
– 24,3 |
17,8±1,7 |
|
|
24,5
– 23,7 |
16,9±1,3 |
|
|
22,1
– 21,6 |
15,7±1,2 |
|
|
20,8
– 19,7 |
14,1±0,9 |
|
|
18,3
– 17,9 |
12,9±1,1 |
|
|
16,0
– 16,6 |
8,3±0,8 |
|
|
14,3
– 13,7 |
3,5±0,3 |
|
|
12,2
– 11,5 |
0,8±0,09 |
|
|
10,2- 10, 4 |
- |
|
|
8,0 |
- |
|
PN растений, обработанных удобрениями, выше PN контрольных
растений во всем диапазоне влажности почвы. В среднем, максимальная скорость PN max наступает при 80% от НВ. Диапазон
PN max (от
90 до 100% относительной скорости фотосинтеза) расширяется в сторону меньших
значений влажности почвы, особенно это заметно в варианте 3. Снижение PN и роста биомассы начинается примерно при 60%
НВ. В области меньших значений
влажности почвы (Рис. 1, точка А) возрастает нефотохимическое тушение,
тенденция к росту темнового дыхания (не показано). Первичная продуктивность
фотосинтеза (фотосинтез минус темновое дыхание) резко снижается, следовательно,
накопление биомассы минимальное. В таблице 1а приведены значения квантового
выхода, транспирации (Tr) и
скорости электронного транспорта (ETR) вариантов
растений при влажности почвы в начале ВЗ (12,0%). Самые высокие значения ETR и Tr при низкой
влажности почвы у растений варианта 3 (с обработкой Микромак и Микроэл). После
проведения измерений растения полили и через некоторое время исследовали их
физиологические характеристики снова (Табл. 1а).
Квантовый
выход ФСII растений
варианта 3 был 0,499 - восстановился до 80% от первоначального, значения ETR max - 139,0 отн. ед., что на 47% ниже
от значений при НВ, нефотохимическое тушение не восстановилось до
прежнего уровня, т.е. высокое. Примерно такие же значения параметров были и у
варианта 3. Восстановление значений ETR
и Tr растений варианта 1, после полива были незначительные, для контрольных растений влажность почвы (10,8%) была близкая к
влажности устойчивого завядания. Для
растений остальных вариантов ВЗ
сдвигалась в сторону меньших значений влажности почвы в диапазоне от 10,8 до
7,8%. По результатам опытов видно, что
растения вариантов с применением удобрений более устойчивые к засухе. Это в первую очередь вследствие
того, что у этих растений хорошо
развита как «надземная часть» так и корневая система. Увеличение биологического азота в почве (как результат ассоциативной
азотфиксации), высокий фотосинтез при небольшом снижении темнового дыхания (в
сравнении с контролем) повышает у
растений устойчивость к засухе.
Таблица 1а. Зависимости квантового выхода (Yield), транспирации (Tr) и скорости электронного транспорта (ETR) от влажности почвы в области наименьшей влагоемкости
(НВ) и влажности завядания (ВЗ) яровой пшеницы Курская 2038 для 3-х вариантов обработки удобрениями.
|
Варианты |
Yield, отн. ед. |
Tr, ммоль/моль |
ETR, отн. ед. |
|||
|
НВ |
около ВЗ |
НВ |
около ВЗ |
НВ |
около ВЗ |
|
|
1 -Контроль |
0,618±0,04 |
0,339 ±0,02 |
3,65 ±0,22 |
0,73 ±0,04 |
120 ±7,2 |
53,0 ±3,2 |
|
2-Микромак |
0,641±0,04 |
0,352 ±0,02 |
3,51 ±0,21 |
1,75 ±0,11 |
119 ±7,1 |
57,2 ±3,4 |
|
3-Микромак +Микроэл |
0,627±0,03 |
0,415 ±0,025 |
5,48 ±0,29 |
2,74 ±0,16 |
128 ±7,7 |
107,6 ±6,4 |
Изменение
холодоустойчивости растений яровой пшеницы при обработке их удобрениями.
Фаза кущение
В исследованиях по реакции растений на
температуры выделены три температурных зоны: зона физиологического комфорта –
фоновая зона (диапазон оптимальных по
фотосинтезу температур), зона физиологического дискомфорта или «зона адаптации
и закаливания», зона повреждающих температур. Для пшеницы эти зоны составляют:
фоновая зона – от 200 до 250С,
низкие температуры зоны адаптации и закаливания – от +50 С до +200
С, и ниже +50С – повреждающая зона. Для исследования
холодоустойчивости мы выбрали температуру воздуха на границе повреждающей зоны и
зоны адаптации (+50 С). При росте растений при температуре воздуха +50 С в течение 8 часов скорость фотосинтеза снизилась примерно в 1,4
- 1,7 раза (Таблица 2). Скорость фотосинтеза PN вариантов
с обработкой удобрениями были 57-60%
для растений варианта 3 от PN растений при
+250 С. В одинаковых условиях роста при +50С PN вариантов с удобрениями была больше PN контроля в 2- 3,8 раза.
По сравнению с контролем ETR
растений вариантов 2 и 3 был больше на
38 и 51%, соответственно. Индекс
жизнеспособности изменялся не так значительно, только в варианте 3 он
увеличился на 9-18%. Зависимости ETR от
интенсивности солнечного излучения показаны на рисунках (Рис. 3, 4).
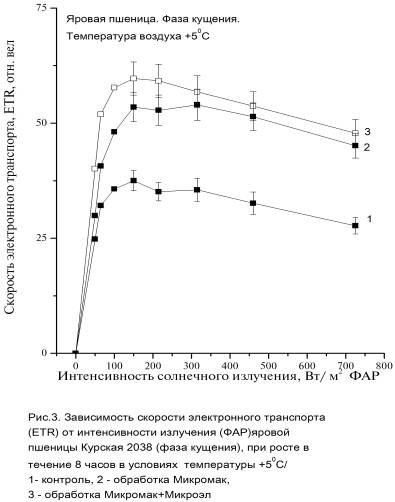
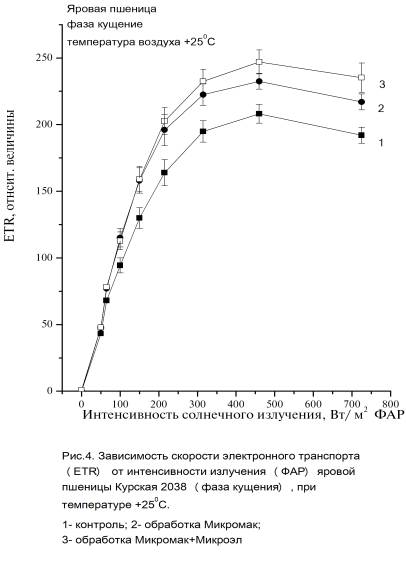
Максимальные
значения ETR max значительно
превышали значения контрольных растений
на 42% для варианта 2 и на 54-59% для варианта 3. При высоких значениях интенсивности
солнечного излучения максимальные скорости электронного транспорта ETRmax вариантов
растений 2 и 3 был значительно выше ETR
контрольных растений, хотя различия между вариантами с обработкой удобрений (варианты
2 и 3) небольшие, т.е. эти растения одинаково реагируют на высокие
интенсивности солнечного излучения.
Снижение PN и ETR
сопровождалось повышением расхода энергии на тепловую диссипацию, повысился
коэффициент нефотохимического тушения (NPQ). Если при температуре воздуха +250 С NPQ было на уровне 0,500 для 1, 2 и
3 вариантов, то при температуре +50 С величины NPQ были 1,706; 1,561;
1,508 соответственно, т.е. увеличились в три раза. В нашем опыте (растения выращивались в сосудах) температура почвы
также снижалась, хотя не так значительно, как воздуха. Это ведет к небольшому снижению
дыхания. Происходит перераспределение
составляющих дыхания в сторону
дыхание поддержания и снижения дыхания роста. Поскольку нефотохимическое тушение в результате повысилось
значительно, то это привело к снижению роста растений и накопления
биомассы.
Растения
варианта 3 имели наибольшие значения по
сравнению с другими PN и ETR и меньшие значения коэффициента нефотохимического
тушения.
Таким образом, варианты растений с
применением Микромак и Микромак +
Микроэл (вар. 3) устойчивее других растений к действию низких температур.
Таблица 2. Влияние температуры воздуха (+50С) на
скорость фотосинтеза (PN),
электронного транспорта (ETR), квантовый
выход фотосистемы II (Yield) и индекс жизнеспособности (IV) листьев растений пшеницы яровая Курская 2038.
Фаза роста «кущение».
|
Варианты удобрений |
PN, мгСО2/
дм2 час |
Yield, отн. вел. |
ETR, отн. вел. |
IV, отн.
вел |
||||
|
+250С |
+50С |
+250С |
+50С |
+250С. |
+50С |
+250С |
+50С |
|
|
Контроль |
7,8 ±0,47 |
2,1 ±0,13 |
0,661 ±0,04 |
0,140 ±0,01 |
102,6 ±6,2 |
29,8±1,7 |
2.94 ±0,18 |
1,47 ±0,09 |
|
Микромак |
9,4 ±0,56 |
4,3±0,26 |
0,639 ±0,04 |
0,222 ±0.01 |
111,7 ±7,0 |
53,5±3,2 |
3,08 ±0,18 |
1,81 ±0,11 |
|
Микромак+ Микроэл |
13,4 ±0,80 |
8,1±0,48 |
0,645 ±0,04 |
0,229 ±0,014 |
118,7 ±7,1 |
55,5±2,8 |
3,2 ±0,19 |
2,07 ±0,12 |
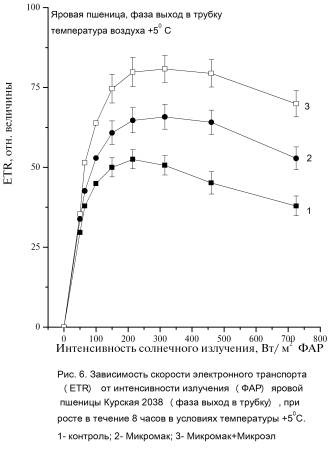
Фаза выход в трубку
В
фазе выход в трубку скорость фотосинтеза PN при
температуре +50С снизилась по сравнению с PN при
температуре +250С на 65%, 47%
и 45% для растений вариантов 1,
2 и 3 соответственно (Таблица. 3). По сравнению с PN контрольных растений (при температуре +50 С) PN растений других вариантов была больше
на 46% - 56% для варианта 2. PN растений 3-го варианта
была в 2,9 – 3 раза больше PN контроля. В вариантах растений с обработкой
удобрениями (при температуре +50С)
величина ETR по
сравнению с контролем увеличилась на 58% для растений вариантов 2. Для растений
3 варианта увеличение ETR составило 82-85%.
Максимальные значения ETR max растений, обработанных удобрениями, превышали значения контрольных растений на 25, 9% для варианта 2 и на 55% для
варианта 3 (Рис. 5, 6).
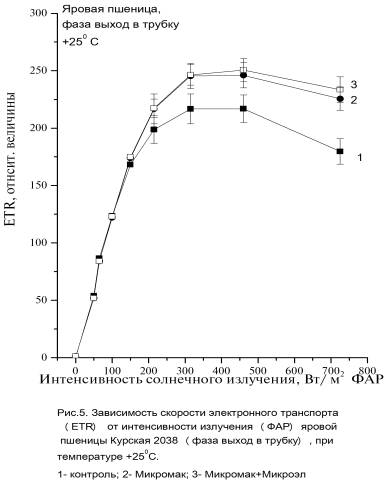
Так
же как и в фазе кущения, у растений при температуре +50С коэффициент нефотохимического тушения вырос
от 0,400 до 1,0 - произошло увеличение
тепловой диссипации в светособирающей антенне ФСII .
Как
в фазе кущения, так и в выход в трубку растения варианта 3 стабильно показывают
высокие значения PN и ETR, У этих
растений также существенно изменилось значение индекса жизнеспособности - увеличилось на 9 и 24% соответственно -
возможно за счет улучшенного азотного
питания и большего содержания хлорофилла в листьях. Таким образом, применение удобрений в целом способствовало адаптации
растений к низким температурам и
закаливанию (т.е. устойчивости растений),
особенно, с комплексным применением Микромак + Микроэл.
Таблица 3. Влияние
температуры воздуха (+50С) на скорость фотосинтеза (PN), электронного транспорта (ETR), квантовый выход фотосистемы II (Yield) и индекс
жизнеспособности (IV) растений пшеницы яровая Курская 2038. Фаза роста
«выход в трубку».
|
Варианты удобрений |
PN, мгСО2/
дм2 час |
Yield, отн. вел. |
ETR, отн. вел. |
IV, отн.
вел |
||||
|
+250С |
+50С |
+250С |
+50С |
+250С. |
+50С |
+250С |
+50С |
|
|
Контроль |
17,4 ±1,2 |
6,1±0,4 |
0,577 ±0,03 |
0,305 ±0,02 |
129,3 ±7,7 |
37,5±22 |
2.94 ±0,18 |
1,76 ±0,10 |
|
Микромак |
18,8 ±1,3 |
8,9±0,5 |
0,639 ±0,04 |
0,326 ±0,02 |
128,7 ±7,7 |
59,5±3,5 |
3,08 ±0,18 |
1,78 ±0,11 |
|
Микромак+ Микроэл |
32,4 ±2,1 |
17,7±1,1 |
0,653 ±0,04 |
0,405 ±0,03 |
128,0 ±7,7 |
69,3±4,1 |
3,2 ±0,19 |
2,18 ±0,13 |
Обработка
растений Триатлоном
В фазе кущения после обработки растений комплексом Триатлон скорость фотосинтеза
снизилась на 15% у растений 1варианта
(Табл. 4). У растений вариантов 2и 3 PN снизилась только на 8% и 10% соответственно. Существенное влияние
обработки Триатлоном на скорость электронного транспорта было у растений варианта 1 – на 7-12%. Значения индекса жизнеспособности снизилось для
растений варианта 1 на 20%, а для варианта
3 – на 15%. Таким образом, в фазе кущения обработка растений Триатлоном
снижает скорость фотосинтеза и электронного транспорта на 10-18% для
растений варианта 1. Наименее
чувствительные к обработке Триатлоном
растения варианта 3.
Таблица 4. Влияние
обработки растений яровой пшеницы «Триатлоном» на скорость фотосинтеза (PN) и электронного транспорта (ETR), квантового выхода фотосистемы II (Yield) и индекса
жизнеспособности (IV). Фаза роста «кущение».
|
Варианты удобрений |
PN, мгСО2/
дм2 час |
Yield, отн. вел. |
ETR, отн. вел. |
IV, отн.
вел |
||||
|
без обраб. |
обработка триатлон |
без обраб. |
обработка триатлон |
без обраб. |
обработка триатлон |
без обраб. |
обработка триатлон |
|
|
Контроль |
7,8±0,5 |
6,6 ±0,4 |
0,652 ±0,04 |
0,518 ±0,03 |
118,0 ±7,1 |
110,0 ±7,2 |
3,65 ±0,22 |
2,94 ±0,18 |
|
Микромак |
9,4±0,6 |
8,7±0,5 |
0,646 ±0,04 |
0,535 ±0,03 |
117,8 ±7,1 |
105,6 ±6,5 |
3,1 ±0,22 |
2,48 ±0,14 |
|
Микромак+ Микроэл |
13,4 ±1,0 |
12,1 ±0,7 |
0,645 ±0,04 |
0,520 ±0,03 |
116,0 ±7,0 |
114,0 ±7,2 |
3,4 ±0,21 |
2,9 ±0,18 |
В
фазе выхода в трубку
действие Триатлона на
физиологические параметры было менее существенным (Таблица 5). Скорость
фотосинтеза существенно не менялась для растений всех вариантов (около 5-7%).
Скорость электронного транспорта ETR снизилась на
13-18% для растений вариантов 2 и 3.
Индекс
жизнеспособности после обработки триатлоном наоборот, немного увеличился (на 10-15%). Это можно объяснить тем, в фазе «выход в трубку» препарат действует в
меньшей степени, чем на более ранних стадиях роста, а также положительными
качествами самого препарата - высокой
селективности по отношению к культуре. Во вторых, небольшая величина дозы Триатлона
может даже повысить некоторые физиологические параметры (это больше
относится к более зрелым растениям - в фазе выхода в трубку). Малая доза
ингибитора может вызывать небольшое
повышение величины физиологического параметра.
Это явление известно, например, при изучении действия поллютантов (сернистый газ) на хвою сосны и ели - мы
наблюдали небольшое повышение скорости фотосинтеза при действии на растения
небольших доз сернистого газа. В нашем случае, при действии Триатлона
повышается индекс жизнеспособности у растений в фазе выхода в трубку.
Таблица 5. Влияние обработки растений яровой пшеницы
«Триатлоном» на скорость фотосинтеза (PN) и электронного транспорта (ETR),
квантового выхода фотосистемы II (Yield) и индекса жизнеспособности (IV).
Фаза
роста «выход в трубку».
|
Варианты удобрений |
PN, мгСО2/
дм2 час |
Yield, отн. вел. |
ETR, отн. вел. |
IV, отн.
вел |
||||
|
Без обраб. |
Обработка триатлон |
Без обраб. |
Обработка триатлон |
Без обраб. |
Обработка триатлон |
Без обраб. |
Обработка триатлон |
|
|
Контроль |
17,4 ±1,2 |
16,7 ±1,2 |
0,577 ±0,04 |
0,543 ±0,03 |
107,0 ±6,4 |
109,4 ±6,5 |
2,94 ±0,18 |
3,0 ±0,19 |
|
Микромак |
18,8 ±1,3 |
17,8±1,3 |
0,646 ±0,04 |
0,535 ±0,03 |
125,0 ±7,6 |
102,7 ±6,2 |
3,1 ±0,19 |
3,48 ±0,20 |
|
Микромак+ Микроэл |
28,4 ±2,1 |
27,8±1,9 |
0,645 ±0,04 |
0,520 ±0,03 |
116,0 ±7,0 |
118,0 ±7,0 |
3,67 ±0,22 |
4,23 ±0,25 |
Влияние
обработки растений удобрениями на устойчивость к фотоингибированию
Снижение активности фотосинтетических
процессов под воздействием избыточного освещения называется
фотоингибированием. При различных видах
стресса избыточный свет, энергия которого не может быть безопасно реализована в
фотосинтетических реакциях, считается одним из основных факторов инактивации
комплексов ФСII. Это снижение может быть
обратимым, вызванным регуляторными механизмами, предотвращающими поступление
энергии возбуждения к реакционным центрам за
счет увеличения тепловой диссипации в светособирающей антенне, и
необратимым (индуцированная светом инактивация комплексов ФСII). Для исследования фотоингибирования мы облучали
часть листа растений высокой интенсивностью света (около 775 Вт/ м2
ФАР) в течение 1 часа. Влияние фотоингибирования исследовали по изменению
отношения Fv/Fm. Уменьшение Fv/Fm
обычно отождествляется с повреждением комплексов ФСII в результате стресса. Обнаружено, что в этих условиях
величина Fv/Fm линейно коррелирует со скоростью выделения кислорода.
Соотношение Fv/Fm
измеряется после темновой адаптации, во время которой регуляторные изменения
релаксируют. В этом случае оставшееся уменьшенное значение Fv/Fm
пропорционально степени необратимой фотоинактивации (фотоповреждения)
комплексов ФСII. Поэтому измерения через
интервалы времени Fv/Fm после обработки позволяют определить часть необратимой
фотоинактивации. После обработки части
листа избыточным светом мы измеряли отношение Fv/Fm
через интервалы времени 1 мин, 5 мин (суммарное: обратимое и необратимое
фотоингибирование) и через 30 мин (необратимое фотоингибирование). Считается, что 20-30 мин темновой адаптации
достаточно для релаксации регуляторных изменений [2].
В фазе кущения самое большое снижение Fv/Fm
было у растений контроля – на 14%, и 18% для ETR (Таблица 6). Fv/Fm варианта 2
снижался не так значительно- на 11%. Для этих растений ETR также снижался
- на 8%. У варианта 3 снижения Fv/Fm и ETR менее существенное (7-10%).
В фазе выход в трубку отношение Fv/Fm и скорость электронного транспорта ETR растений
варианта 3 уменьшились на 7- 5% ,что находится в диапазоне погрешностей
(Таблица 7). Изменение этих параметров
для варианта 1 наиболее существенное – от 13 до 17%. Для варианта 2 снижение величин измеряемых параметров находились в
пределах 10%.
По результатам исследования можно сделать вывод, что
действие избытка света более существенное для контрольных растений. Для растений вариантов с обработкой Микромак + Микроэл (вар. 3) изменения Fv/Fm и ETR были не существенные.
Таблица 6. Влияние фотоингибирования (действия высокой
освещенности*) на скорость электронного транспорта (ETR) и максимального квантового выхода фотосистемы II (Fv/Fm) растений яровой пшеницы Курская 2038. Фаза «кущение»
|
Варианты удобрений |
Максимальный квантовый выход фотосистемы II**, Fv/ Fm, отн. велич. |
ETR, отн. велич. |
||||
|
До облуч. |
1 мин после облучения |
5 мин после облуч. |
30 мин после облуч. |
до облуч. |
30 мин после облуч. |
|
|
Контроль |
0,779 ±0,05 |
0,553 ±0,03 |
0,654 ±0,04 |
0,672 ±0,04 |
128,7 ±7,7 |
105.5 ±6,3 |
|
Микромак |
0,788 ±0,05 |
0,614 ±0,04 |
0,693 ±0.05 |
0,701 ±0,04 |
115,8 ±7,0 |
106,8 ±7,5 |
|
Микромак+ Микроэл |
0,798 ±0,05 |
0,630 ±0,05 |
0,686 ±0,05 |
0,742 ±0,05 |
127,7 ±7,7 |
115,5 ±7,9 |
Примечание:
*
- часть листа растения каждого варианта
в течение часа облучали с интенсивностью света 775 Вт/ м2
ФАР.
**
- измерения Fv/Fm
проведены в темноте через 1 мин, 5 и 30 мин после выключения ингибирующего
света.
Таблица 7. Влияние фотоингибирования (действия высокой
освещенности*) на скорость электронного
транспорта (ETR) и максимального квантового
выхода фотосистемы II (Fv/Fm)
растений яровой пшеницы Курская 2038.
Фаза роста «выход в трубку».
|
Варианты удобрений |
Максимальный квантовый выход фотосистемы II**, Fv/ Fm, отн. велич.
|
ETR, отн. велич. |
||||
|
До облуч. |
1 мин после облучения |
5 мин после облуч. |
30 мин после облуч. |
до облуч. |
30 мин после облуч. |
|
|
Контроль |
0,753 ±0,05 |
0,570 ±0,03 |
0,674 ±0,04 |
0,655 ±0,04 |
161,7 ±9,7 |
134.1 ±8,7 |
|
Микромак |
0,756 ±0,05 |
0,604 ±0,04 |
0,665 ±0,04 |
0,672 ±0,04 |
182,5 ±10,7 |
166,2 ±9,7 |
|
Микромак+ Микроэл |
0,779 ±0,05 |
0,665 ±0,04 |
0,704 ±0,05 |
0.732 ±0,05 |
187,7 ±10,7 |
178,6 ±9,9 |
Примечание:
*
- часть листа растения каждого варианта в течение часа облучали с интенсивностью света 775 Вт/ м2
ФАР.
**
- измерения Fv/Fm проведены в темноте через 1 мин,
5 и 30 мин после выключения ингибирующего света.
Выводы
1.
Засухоустойчивость.
По
результатам опытов видно, что растения вариантов с применением
удобрений
более устойчивые к засухе. Наиболее
устойчивы к низким значениям влажности почвы варианты с обработкой семян
Микромак и некорневыми обработками Микроэл.
Комплексная обработка удобрениями
увеличивает рост растений, фотосинтез в 1,8 раза, в 3 раза увеличивается
азотфиксация в почве. Известно, что при хорошем азотном питании повышается
устойчивость растений к засухе.
2.
Холодоустойчивость.
При росте в ранней фазе кущения лучшие
растения по холодоустойчивости были при обработке Микромак и некорневой обработкой Микроэл. При
температуре +50С PN вышеуказанных вариантов была больше PN
контроля в 2- 3,8 раза, в среднем фотосинтетическая активность этих растений была в 1,5-1,8 раза выше, чем в контроле.
В
фазе выхода в трубку устойчивее других
растений к действию низких
температур были растения с комплексной
обработкой Микромак и некорневой обработкой Микроэл. Скорость фотосинтеза (PN) растений
этого варианта были в 2,9 – 3 раза больше PN контроля.
3.
Обработка растений
Триатлоном.
Триатлон влияет на растения пшеницы в
фазу кущения, т.е. в фазу, рекомендуемую для химпрополки. В этой фазе
роста обработка растений Триатлоном
снижает скорость фотосинтеза и электронного транспорта на 10-18% для растений
вариантов контроль. У растений совместной обработки семян Микромак и некорневой
обработки Микроэл воздействие Триатлона на фотосинтетическую деятельность было
минимальное
В фазе выхода в трубку Триатлон не оказывал действия на фотосинтетическую активность растений.
Скорость фотосинтеза существенно не менялась для растений всех вариантов (около
5-7%). В целом, для обеих фаз роста растений устойчивее к действию Триатлона
были растения с комплексной обработкой удобрениями (вар. 3).
4. Фотоингибирование.
При избытке
света в фазе кущения величины скорости фотосинтеза и других показателей
фотосинтетической активности растений у
контрольных растений снижались в среднем на 15%, у варианта с обработкой
только Микромак это снижение составляло
около 10%. У растений вариантов с обработкой
Микромак + Микроэл изменения
физиологических параметров были в пределах погрешности измерений, т.е. эти
растения были наиболее устойчивы к фотоингибированию.
В целом, растения вариантов с обработкой
семян Микромак и некорневой обработкой
Микроэл имеют хорошо развитую как
«надземную часть» так и корневую систему, высокий уровень хлорофилла, высокую
скорость фотосинтеза, высокий коэффициент использования солнечной энергии на
фотосинтез (по измеренным величинам квантового выхода растений). Все это, а
также увеличение биологического азота в почве (как результат ассоциативной
азотфиксации) значительно повышает у растений
устойчивость к исследованным стресс-факторам.
ЛИТЕРАТУРА
1. Мудрик В.А., Куликов С.С., Новичкова Н.С., Редькина
Н.В., Лучицкая О.А., Иванов Б.Н. Влияние микроудобрений на ассоциативную
азотфиксацию и продуктивность яровой пшеницы. Агрохимия. 2010. № 9, с. 72-76.
2. Корнеев Д.Ю. Информационные возможности метода
индукции флуоресценции хлорофилла. Киев: Альтпресс. 2002. 188 с.
3. H.K. Lichtenthaler,
C. Buschmann, and M. Knapp. How to correctly determine the different chlorophyll
fluorescence parameters and chlorophyll fluorescence decrease ratio Rfd
of leaves with the PAM fluorometer. Photosynthetica. 2005. 43(3), pp 379-393.