Биологические
науки / 2. Структурная ботаника и биохимия растений
К.б.н. Шуляковская Т.А., к.б.н. Ильинова М.К.
Институт леса Карельского научного центра Российской
академии наук, Россия
Особенности липидного состава тканей ствола Betula pendula Roth при образовании узорчатой древесины
Betula pendula var. carelica (Mercklin) Hämet-Ahti – карельская береза – ценное и морфологически необычное
растение. Она является формой вида Betula pendula Roth. От другой формы данного вида
– березы повислой Betula pendula var. pendula – карельская
береза отличается аномальной узорчатой
древесиной. Своеобразные темно-коричневые штрихи в сочетании с дающей блеск
свилеватостью приводят к возникновению на обработанных поверхностях изделий из
карельской березы неповторимого мраморовидного рисунка. Наибольшую
хозяйственную ценность имеет сильно свилеватая древесина с многочисленными
коричневыми включениями, которые представляют собой аномальные скопления
паренхимы (Коровин и др., 2003).
В нашей работе саженцы карельской березы из
сертифицированных семян от контролируемого опыления, закупленных в Финляндии в
фирме Forelia OY.,
растущие в одинаковых условиях, к семилетнему возрасту имели разные особенности
внешнего и внутреннего строения ствола. Среди них мы выделили две группы растений:
саженцы нормального строения (ровные гладкие стволы, прямослойная древесина) и
саженцы аномального строения, что внешне выражалось в образовании на стволах
утолщений, а также изменении текстуры древесины ствола, появлении узорчатости
древесины. Для раскрытия механизма такого развития необходимо изучить
особенности метаболизма, определяющие структурные отклонения от нормы. Целью
данной работы было исследование содержания и жирнокислотного состава липидов,
сравнительная характеристика данных показателей у саженцев с нормальным и
аномальным развитием ствола. Это поможет выделить особенности липидного
метаболизма, характерные для формирования аномальной структуры древесины
карельской березы.
У карельской березы с аномальным развитием
(проявившейся) ствол (ниже кроны) разделили на 4 части. Самая верхняя часть
ствола имела слабое проявление узорчатости древесины, самая нижняя была
наиболее насыщена по рисунку древесины. Сверху вниз по стволу наблюдалось
усиление насыщенности рисунка древесины, т.е. признака «карелистости». У
карельской березы нормального развития (непроявившейся) отбирали образцы тканей
из верхней, средней и нижней части ствола. Проводили отбор феллодермы –
паренхимы коры и наружного слоя древесины. Отбор образцов (спиливание саженцев)
проходил в период активной камбиальной деятельности (утолщения ствола) – вторая
половина июля (возраст саженцев 7 лет) и во время покоя по окончании
вегетационного периода – вторая половина октября 2009 г., когда саженцы
становились уже восьмилетними. Растительный материал фиксировали в жидком
азоте, высушивали в лиофильной сушилке. Экстракцию липидов проводили смесью
хлороформа и метанола в соотношении 2:1 по объему. Фракционирование суммарных
липидов осуществляли методом колоночной хроматографии. На колонку с силикагелем
Bio-Sil A100-200 mesh переносили липидный экстракт, растворенный в
хлороформе, и элюировали растворителями отдельные фракции: хлороформом –
нейтральные липиды (НЛ), ацетоном – гликолипиды (ГЛ), метанолом – фосфолипиды
(ФЛ). Жирные кислоты (ЖК) липидных фракций исследовали в виде их метиловых
эфиров, которые получали переэтерификацией липидов метанолом в присутствии
ацетилхлорида, и разделяли на хроматографе «Хроматэк – Кристалл-5000.1» (Йошкар-Ола,
Республика Марий Эл). Колонка Zebron
ZB-FFAP капиллярная
50м х 0,32мм. Идентифицировали жирные кислоты с помощью метчиков – стандартных
жирных кислот (Supelco 37 компонентов). Содержание
жирных кислот в липидах определяли с помощью внутреннего стандарта –
маргариновой кислоты, которую вносили в определенном количестве в образец перед
метилированием. Все жирные кислоты распределяли по группам в зависимости от
степени ненасыщенности: моноеновые (М) – в углеродной цепочке имеется одна
двойная связь; диеновые (Д) – две; триеновые (Тр.) – три; тетраеновые (Тетр.) –
четыре и насыщенные (Н) – без двойных связей. Индекс 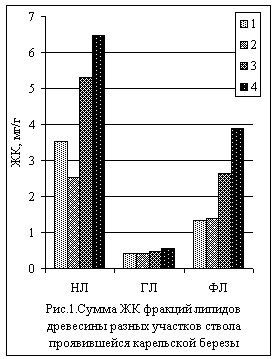 ненасыщенности (ИН) рассчитывали по методу D. Lyons:
ненасыщенности (ИН) рассчитывали по методу D. Lyons:
ИН=(1М+2Д+3Тр.+4Тетр.)/100
Проведенные исследования показали, что у проявившейся карельской березы в направлении увеличения узорчатости древесины, т.е. сверху вниз по стволу, росло содержание жирных кислот (а, следовательно, и липидов) всех трех липидных фракций древесины. Количество нейтральных липидов возрастало в 2,5 раза, гликолипидов – в 1,3 раза, а фосфолипидов – наиболее существенно: в 2,9 раза (рис.1).
При сравнении содержания жирных кислот в
древесине и феллодерме одних и тех же участков ствола в июле отмечено, что в
верхней половине ствола (1-2 участки) имело место преобладание жирных кислот
нейтральных липидов в феллодерме в 3,6 – 3,8 раза, а к участкам 3-4 наступало
преимущество древесины по содержанию жирных кислот нейтральных липидов в 1,2 –
1,4 раза. В верхней части ствола (сразу под кроной) в паренхимных клетках
флоэмы накапливались ассимиляты, образованные в листьях в результате активного
фотосинтеза и не использованные в процессе роста, их излишек превращался в
липиды и оседал в паренхимных клетках (в феллодерме). В нижних участках ствола
таких запасов нейтральных липидов в паренхиме флоэмы становилось меньше, но
росло количество подобных запасов в ксилеме (древесине). Чем ниже по стволу
участок древесины, тем выше запасы нейтральных липидов в ней и тем более
узорчатый рисунок у древесины. Накопление запасных липидов в паренхимных
клетках древесины сопровождалось повышением свилеватости транспортных путей и
усилением узорчатости древесины карельской березы.
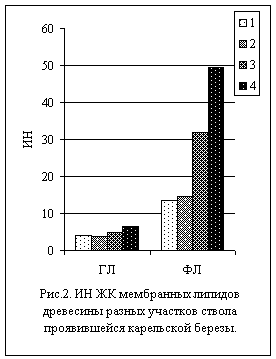
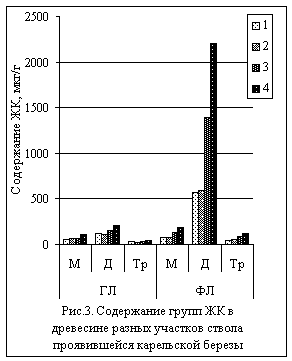
В фосфолипидах древесины сверху вниз
существенно рос индекс ненасыщенности (в 3,6 раза) (рис.2), главным образом, за
счет диеновых кислот, которые увеличивали свое содержание в 4 раза к нижнему
участку ствола (рис.3). В основном, это линолевая кислота (18:2). У
гликолипидов при этом индекс ненасыщенности возрастал в 1,6 раза, а диеновые
кислоты (в основном, линолевая) – в 1,7 раза. Таким образом, в участках
древесины ствола, содержащих много нейтральных липидов, существенно возрастал
уровень фосфолипидов, причем среди жирнокислотных остатков этих фосфолипидов
преобладала линолевая кислота. Значит, повышенное образование липидных капель в
узорчатой древесине приводило к увеличению содержания мембранных липидов
(фосфолипидов), а последние нарастали за счет липидов с высоким содержанием линолевой
кислоты. Дополнительный синтез фосфолипидов мог быть связан с необходимостью
образования дополнительных мембран, ограничивающих многочисленные липидные
капли.
Главная функция мембранных липидов состоит
в том, что они формируют бислойный матрикс, с которым взаимодействуют белки. От
состава мембранных липидов, то есть от наличия двойных связей, или от степени
ненасыщенности жирнокислотных остатков, зависит плотность упаковки липидов и,
соответственно, текучесть мембраны. Физическое состояние липидного бислоя
влияет на каталитическую активность многих мембранных ферментов (Геннис, 1997;
Гринштейн, Кост, 2001). Фосфолипидное окружение ферментного белка в мембране
играет важную роль в поддержании определенной конформации фермента, а значит, и
его активности, или служит для облегчения взаимодействия фермента и субстрата.
Молекулярное строение фосфолипидов позволяет без каких-либо изменений структуры
молекулы, лишь заменой входящих в состав липида жирных кислот с присущими им
качествами менять физико-химические свойства липида, позволяет влиять на
функциональные характеристики мембран и на условия деятельности мембранных
белков, не затрагивая белковую часть мембраны (Крепс, 1981).
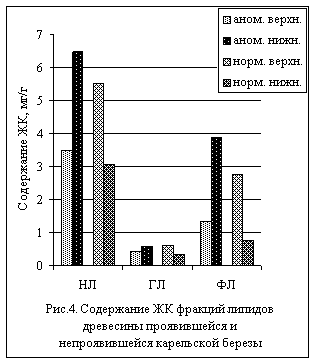 Сравнение проявившейся и непроявившейся карельских
берез между собой показывает, что в древесине первой из берез содержание ЖК НЛ
и ФЛ значительно преобладает в нижней части ствола по сравнению с верхней,
тогда как у непроявившейся березы картина обратная, а именно, заметное
преобладание отмечено в верхней части ствола (рис.4). Т.е. наблюдается
зеркальное отражение соотношения уровней ЖК липидных фракций в древесине разных
частей ствола у берез с разной текстурой древесины.
Сравнение проявившейся и непроявившейся карельских
берез между собой показывает, что в древесине первой из берез содержание ЖК НЛ
и ФЛ значительно преобладает в нижней части ствола по сравнению с верхней,
тогда как у непроявившейся березы картина обратная, а именно, заметное
преобладание отмечено в верхней части ствола (рис.4). Т.е. наблюдается
зеркальное отражение соотношения уровней ЖК липидных фракций в древесине разных
частей ствола у берез с разной текстурой древесины.
В результате вегетационного периода происходило
накопление нейтральных (можно считать, запасных) липидов в наиболее узорчатой
части древесины ствола карельской березы. В древесине проявившейся карельской
березы содержание ЖК мембранных липидов и их ненасыщенность росли одновременно,
т.е. рост ЖК липидных фракций происходил за счет ненасыщенных кислот.
Выводы:
•Узорчатая древесина карельской березы
содержит в клетках паренхимы значительное количество нейтральных липидов,
которые в большинстве своем представляют запасные липиды.
•При этом в узорчатой древесине отмечен
существенный рост содержания и ненасыщенности жирных кислот фосфолипидов, в
основном, за счет линолевой кислоты.
•Увеличение ненасыщенности жирных кислот фосфолипидов
указывает на изменение свойств клеточных мембран, а, следовательно, и их
функционального состояния, что приводит к изменению метаболизма и
направленности дифференциации клеток камбия ствола карельской березы.
•Сравнение проявившейся карельской березы
с непроявившейся показывает противоположный характер изменения содержания
липидных фракций в древесине по длине ствола: у карельской березы с признаками
«карелистости» содержание всех трех фракций липидов растет к нижней (наиболее
узорчатой) части ствола, а у березы без признаков падает.
Литература:
Геннис Р. Биомембраны: Молекулярная структура и функции. М.,
1997. 622 с.
Гринштейн
С.В., Кост О.А. Структурно-
функциональные особенности
мембранных белков // Успехи биологической химии. 2001.
Т. 41. С. 77-104.
Коровин В.В.,
Новицкая Л.Л., Курносов Г.А.
Структурные аномалии стебля древесных растений. М., 2003. 280 с.
Крепс Е.М. Липиды
клеточных мембран Л., 1981. 339 с.