Біологічні науки/Структурна ботаніка і біохімія
Пюрко О.Є.,
Галаган Т.Ж.
Мелітопольський державний
педагогічний університет імені Б.
Хмельницького
Структурна специфічність фотосинтезуючих органів рослин
північно-західного Приазов’я
Формування флори північно – західного Приазов’я
пояснюється регіональними кліматичними умовами, а також екологічними
показниками – зволоженістю, кислотністю, трофністю, вмістом у грунті надлишку
солей. Все більше уваги приділяється вивченню галофітів, так як засолені землі
на Україні займають великі площі (76 тис. га) [3,4]. Засоленою являється
приблизно 25% всієї землі планети. Вони займають великі території в Середній
Азії, в пустинях Африки, прибережних морських районах всіх країн. Приазов’я
розташоване в степовій зоні. Засоленість грунтів узбережжя зумовлюють наявність
морської акваторії (Азовське море, Молочний лиман) та інтенсивне випаровування
води. Вміст солей у грунті визначає процеси життєдіяльності організму на всіх
рівнях його організації, починаючи з молекулярного і закінчуючи фітоценозом і
являється для рослин екзогенним фактором середовища [1,2]. Збільшення вмісту
солей в свою чергу приводить до зниження врожайності сільськогосподарських
культур. Актуальною проблемою на сьогодення є вивчення анатомічної біорізноманітності
рослин північно – західного Приазов’я.
Наші дослідження довели, що листки Halimione verrucifera L. дуже дрібні, овальні до продовгувато – яйцевидних,
цільнокрайні, на поперечному розрізі округлі, продовгуваті. Епідерма
розташована в два шари клітин. Зверху знаходяться великі овальні клітини.
Функція епідерми – захищати листок від висихання, механічних дій. Епідерма
складається з основних епідермальних клітин і продихових. Основні клітини –
безкольорові, характеризуються певною формою та розмірами. У Halimione verrucifera L. виділяють верхній і нижній епідерміс листку.
Дослідження
показали, що верхній епідерміс листку Halimione verrucifera L. має клітини з прямолінійними обрисами, округлою
проекцією, кути - закруглені та загострені, кількість на 1 мм2 – 110
шт., коротка вісь – 1,5 мкм; розмір по довгій вісі – 214,5 мкм; площа клітини –
311 мм2 . Нижній епідерміс має прямокутну проекцію площі
епідермальних клітин, обриси прямолінійні, кути в суміжних межах загострені та
закруглені, кількість клітин на 1 мм2 – 130шт., розмір по довгій
вісі – 31,7 мкм, по короткій – 25,5 мкм; площа клітини – 808,4 мм2 .
Продихи аномоцитного типу, їх кількість на адаксіальній стороні складає 75-80
шт/мм2, а на абаксіальній – 85-90 шт/мм2 .
Таким чином,
наші дослідження показали, що листок вкритий з адаксіальної та абаксіальної
сторін епідермою. Продихи аномоцитного типу. За внутрішньою будовою листок –
атріплекоїдного типу, має кранц-анатомію. Провідна система знаходиться
всередині водоносної паренхіми, але закінчення її виходять на периферію та
лежить в контакті з клітинами хлоренхімної обкладки. Співвідношення тканин
листку приведено в рис. 1.
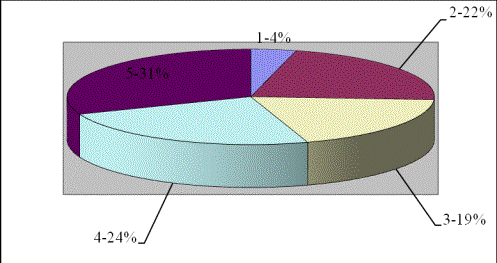
Рис. 1. Співвідношення тканин
листку Halimione verrucifera L.: 1 – епідерма, 2 - фотосинтезуюча
паренхіма, 3 - водоносна паренхіма, 4 – хлоренхіма. 5 – СВП.
Гістологічне
вивчення листку Taraxacum bessarabicum довело, що зверху він вкритий кутикулою (8![]() 2,3 мкм). Епідерма листку включає три типи основних клітин на
абаксіальній поверхні:
2,3 мкм). Епідерма листку включає три типи основних клітин на
абаксіальній поверхні:
I тип –
клітини неправильної форми; проекція їх розпластана, обриси зигзагообразні,
кути в суміжних межах гострі, кількість на 1мм2 - 75 шт., розмір по
довгій вісі - 92,3 мкм, по короткій - 57 мкм, S=5261,1 мкм2.
ІІ тип –
прозенхімні клітини; проекція яких витягнута, обриси прямолінійні, кути в
суміжних межах гострі, кількість на 1 мм2 - 55 шт.; розмір по довгій
вісі - 100 мкм; по короткій - 35,4 мкм; S=3540 мкм2.
III тип –
паренхімні клітини з округлою проекцією та обрисами, кути в суміжних межах
тупі, кількість на 1мм2 - 10 шт.; розмір по довгій вісі - 41,3 мкм;
по короткій - 38 мкм; S=1569,4 мкм2.
Адаксіальна
епідерма листку має такі типи клітин:
I тип –
прозенхімні клітини з витягнутою
проекцією; обриси яких прямолінійні; кути в суміжних межах гострі; кількість на
1 мм2 - 34 шт.; розмір по довгій вісі - 107 мкм; по короткій - 38,6
мкм; S=4130,2 мкм2.
ІІ тип –
клітини паренхімні з округлою проекцією та
обрисами; кути в суміжних межах закруглені; кількість клітин на 1 мм2
- 33 шт.; розмір по довгій вісі - 56,5 мкм; по короткій - 55 мкм; S=3107,5 мкм2.
III тип –
клітини неправильної форми; проекція - розпластана, обриси зигзагообразні, кути
в суміжних межах гострі, кількість на 1 мм2 - 61 шт.; розмір по
довгій вісі 97,5 мкм; по короткій - 82 мкм; S=7995 мкм2. В епідермі
знаходяться великі моторні клітини, котрі мають тонку обкладку і при підсиханні
скручують лист. У Taraxacum bessarabicum амфістоматичний тип листку. Тип продихових комплексів –
аномоцитний; на абаксіальній їх 115 шт/мм2, на адаксіальній – 120
шт/мм2.
Вивчення
внутрішньої будови листку цього виду визначило, що епідерма складає 8% від
загальної товщини листку, під нею знаходиться гіподерма, розміри якої 7%.
Клітини гіподерми округлої форми, які близько прилягають одна до одної. Під
гіподермою знаходиться хлоренхіма (83%), яка складається зі стовбчастого та
губчатого мезофілу, з клітинами округлої форми, розташованими окремо одна від
одної. СВП – закритого, колатерального типу. Співвідношення тканин листку
приведено в рис. 2.
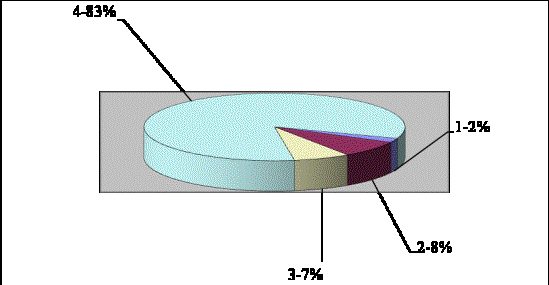
Рис.
2. Співвідношення тканин листку Taraxacum bessarabicum: 1 – кутикула, 2 – епідерма, 3 – гіподерма, 4 - хлоренхіма
Виходячи с
цього, можна зробити наступні висновки:
1.
Структурний аналіз вегетативних органів рослин дозволяє стверджувати, що листок
Halimione verrucifera L. амфістоматичний, продихові комплекси аномоцитного типу.
Має атриплекоїдний тип кранц – анатомії. Halimione verrucifera L. відноситься до
ксерогалофіту, являється субдомінантним в галофітних фітоценозах
північно-західного Приазов’я.
2. У Taraxacum bessarabicum – амфістоматичний тип
листку. Тип продихових комплексів аномоцитний. Мезогалофітного типу кранц –
анатомії. За кількісно – анатомічними особливостями вегетативних органів Taraxacum bessarabicum можна віднести до
мезогалофіту, гемікріптофіту.
Література
1.
Баранова М.А. Классификация морфологических типов устьиц //Ботанический журнал. – 1985. - Т. 70,
№12. - С. 1585 –
1595.
2. Вознесенская Е.В., Гамалей Ю.В.
Ультраструктурная характеристика листьев с кранц – анатомией //Бот. журнал, 1986. – Т.71,
№10. – С. 1291 – 1305.
3. Дідух Я.П.,
Шеляг – Сосонко Ю.Р. Класифікація екосистем – імператив національної екомережі
(ECONET) України //Укр. ботан. журн., – 2001. - № 4. – С. 393 – 403.
4. Савін В. Збереження
ландшафтного та біологічного різноманіття у Запорізькій області //В кн.
Збереження та моніторинг біол. та ланд. різноманіття в Укр. - К., 2000. –
С.110.