Свистунова В.В., Гаврилюк В.Г.,
Пилявська О.А., Вінніков А.І.
Дніпропетровський національний
університет, біолого-екологічний факультет, кафедра мікробіології і
вірусології.
Морфолого-культуральні та біологічні властивості
ізолятів збудників грибкових захворювань зернових культур
На озимій пшениці паразитують понад 10 видів
фітопатогенних грибів родини Septoria, серед яких найпоширеніші та
найшкідливіші Septoria tritici та
Septoria nodorum. Для обмеження септоріозу важливою умовою є правильна
діагностика захворювання. Вона ускладнюється тим, що всихання листя під дією
патогена часто приймають за їх нормальне фізіологічне старіння. Тому, було
проведено дослідження по розробці та удосконаленню методів виділення,
культивування та вивчення впливу фітопатогенних грибів на процеси розвитку та
врожайності зернових культур. Спостереженнями за розвитком культур Septoria tritici на
живильних середовищах виявили два типи: дріжджеподібний й міцеліальний.
Дріжджеподібні колонії майже цілком складалися із вторинних конідій, що
утворюються на зразок бруньок. Тому в дріжджеподібній культурі одночасно
перебували спори різної морфології. Лише іноді в колоніях розвивався вбогий
міцелій. Колонії міцеліального типу не представляли відхилень від норми. Колонії S.tritici відрізнялися
повільним ростом. При цьому міцеліальні ізоляти розвивалися швидше, ніж
дріжджеподібні.
Так, у
міцеліальних ізолятів 18, що швидко ростуть на крохмально-глюкозному агарі
(КГА) й пептонно-сольовому агарі (АПС), 7 й 10 на КГА діаметр колоній через 30
діб рівнявся відповідно 43,3 й 39,2-39,3 мм, тоді як діаметр найбільшої
дріжджеподібної колонії 31 на КГА й АПС склав через такий же проміжок часу 33,9
й 30,3 мм. Відзначено істотні розходження в розмірах колоній і серед ізолятів
одного типу. Так, на КГА дріжджеподібні ізоляти 3 й 31 мали відповідно колонії
діаметром 20,9 й 33,9 мм; у міцеліальних ізолятів 6 й 7 на КГА ці параметри
рівнялися 28,5 й 39,2 мм відповідно.
Швидкість росту
колоній окремих ізолятів залежала від складу живильного середовища. Найбільш
сприятливими для росту майже всіх ізолятів S.tritici являються КГА й АПС. На
останній трохи гірше, ніж на КГА, росли ізоляти 7, 10, 31 й 32. На середовищі
Чапека в більшості випадків розвиток гриба затримувався, розмір колоній тут був
іноді майже в 1,5-2 рази менше, ніж на інших середовищах (15,5 й 29,0 мм в
ізолятах 16 й 18 на середовищі Чапека й 29,2 й 43,3 мм у цих же ізолятах
відповідно на АПС). На середовищі Чапека не спостерігалось ні підвищення, ні
пригнічення розвитку ізолятів 3, 7, 10, і 32, а в ізоляті 19 діаметр колонії на
цьому середовищі був більше, ніж на двох інші.
Ізоляти
S.tritici значно розрізнялися між собою по споруляції. У всіх варіантах досліду
перевага завжди залишалася за ізолятами дріжджеподібного типу, які продукували
на однім і тім же середовищі майже в 30, а іноді в 300 разів більше спор, ніж
міцеліальні; так, було виявлено 444,2 млн. спор в дріжджеподібному ізоляті 16 у
порівнянні з 16,8 млн. спор в міцеліальному ізоляті 21 на АПС, а також 58,2 млн. спор в дріжджеподібному
ізоляті 17 на середовищі Чапека в порівнянні з 0,2 млн. спор в міцеліальному
ізоляті 21 на цьому ж середовищі. Інтенсивність спороносіння окремих ізолятів
залежала від складу живильного середовища. На середовищі АПС завжди відмічалась
найбільша інтенсивність споруляції: до 1,5, а іноді до 80 разів вихід спор того
або іншого ізолята був вищий в порівнянні з іншими субстратами. На середовищі
Чапека спостерігалась затримка споруляції більшості ізолятів що вивчалися, і
лише в деяких ізолятах (16, 17 й 18) число спор на цьому середовищі було
однаковим із КГА. Зовнішній вигляд колоній ізолятів що випробовувалися на КГА й
АПС був, як правило, однаковим. Колонії дріжджеподібного типу слизові, вологі,
блискучі; поверхня піднята й дуже гофрована; край колоній хвилястий, нерівний.
Колонії можуть бути пофарбовані цілком у блідо-рожевий або чорний колір або
мати темну середину й блідо-рожевий край. Колонії міцеліального типу з добре
розвиненим міцелієм. Округлі, часто з нерівними краями; поверхня іноді складчаста;
колір від білого до темно-сірого. На середовищі Чапека всі ізоляти формували
колонії з дуже рідким, сланким міцелієм. Результати експериментів по вивченню
морфолого-культуральних властивостей ізолятів Septoria nodorum
показали, що досліджувані ізоляти на різних живильних середовищах розвивалися
неоднаково. Колонії ізолятів 1 й 4 значно відставали в рості при культивуванні
на АПС. Повільний розвиток на АПС і на середовищі Чапека був у ізолята 8. Для
ізолята 23, навпаки, АПС виявився самим сприятливим субстратом, а найменш
підходящим - картопляно-глюкозний агар. Ізоляти 5, 12, 14, 15 й 22 однаково
добре росли на всіх живильних середовищах. При вирощуванні на одному й тому ж
живильному середовищі швидкість росту ізолятів значно варіювала. Всі ізоляти,
за винятком ізолята 5, мали колонії з добре розвиненим порошистим, шерстистим
або бархатистим міцелієм. У ізолята 5 на КГА міцелій розвивався слабко; колонія
була гранульованою через наявність великої кількості пікнід. Колір колоній
ізолятів на живильних середовищах змінювався від біло-рожевого або
жовтувато-білого до сірувато-зеленого й темно-маслинового. У ізолята 12 на КГА,
ізолятів 1, 4, 5, 12, 14 й 23 на середовищі Чапека й майже у всіх ізолятів
(крім ізолятів 5 й 12) на АПС відзначалася по-різному виражена складчастість
колоній. Ізоляти також розрізнялися між собою по інтенсивності спороносіння.
Високоспорулюючими були ізоляти 1, 4, 5, 12 й 14, інтенсивність спороносіння
яких становила 15-33 млн. спор на 1 см2 живильного субстрату. Набагато слабкіше
спорулювали ізоляти 15 й 23 – біля 1- 4 млн. спор на 1 см2 живильного субстрату. Дуже слабко спорулювали
ізоляти 8 й 22 – до 200-500 тис. спор/см2.
На споруляцію
окремих ізолятів впливав склад живильного середовища. Наприклад, на
пептонно-сахарозному агарі в ізолятів 4 й 14 в 6-8 разів був збільшений вихід
спор у порівнянні із КГА й середовищем Чапека. Остання пригнічувала утворення
спор в ізолятів 1, 4, 12 й 23; кількість їх було в 2, а в окремих ізолятів в 80
разів менше, ніж на КГА й АПС. Так, ізолят 12 на 1 см2 середовища
Чапека формував усього 190 тис. спор, тоді як на КГА й АПС число їх на цій же
площі коливалося від 11 до 17 млн. На середовищі не спостерігалось ні
підвищення, ні пригнічення інтенсивності спороносіння ізолятів 8, 15 й 22. На
наступному етапі були проведені дослідження по вивченню патогенетичного впливу
виділених ізолятів фітопатогенних грибів Septoria tritici i Septoria nodorum на
процеси розвитку і показники врожайності різних сортів озимої та ярової
пшениці, що вирощують у Дніпровському й Солонянському районах Дніпропетровської
області. Ізоляти S.tritici відрізнялися між собою по ступеню патогенності.
П'ять ізолятів були високопатогенними й вражали більш ніж на 50% по 9-11 сортів
пшениці, при цьому у двох інших сортів вони викликали середній ступінь поразки.
Меншу патогенність проявляв ізолят 7, що вражав у сильному ступені (на
55,0-81,7 %) п'ять й у середньому ступені - шість сортів. Ізолят 3 вражав всі
сорти в середньому ступені - від 17,0 до 45,9 %. Всі сорти, що брали участь у досліді, були
высокочутливими до S.tritici і уражувалися в значній мірі (на 50,5-100 %)
чотирма - шістьома ізолятами збудників. В окремих випадках спостерігалася
кореляція між вірулентністю й інтенсивністю споруляції гриба in vitro. Так,
ізолят 16, що вражав у значній мірі майже всі досліджувані сорти, що
випробовувалися, формував на них і велику кількість пікнід - до 350 на одному
листі (за виключенням сортів Харківська 46 й Одеська 51, на яких кількість
пікнід на одному листі рівнялася 15 й 47 відповідно). Такий же зв'язок можна
відзначити для ізолятів 3, 18 й 21. Не відзначено прямої кореляції між
зазначеними ознаками тільки для двох ізолятів 17 й 19. Очевидно, ізолят можна
вважати більш патогенним, а сорт більше сприйнятливим, якщо високий ступінь
поразки сорту сполучається з високою інтенсивністю споруляциії гриба на ньому,
і навпаки. При порівнянні цих двох ознак більшу стійкість постійно показували
озимий сорт Одеська 51 й яровий Харківська 46, на яких приблизно при однаковій
з іншими сортами середній ураженості спостерігалася слабка споруляція (у
середньому близько 19 й 23 пікнід на одному листі). На більш сприйнятливих
сортах Безоста і Донецька 74 при майже рівній з вищевказаними сортами середній
ураженості листів формувалося на одному листі близько 95 й 137 пікнід
відповідно. Із усіх випробуваних ізолятів, найбільш патогенними виявилися
чотири – 16, 17, 19 і 21 (рис.1).
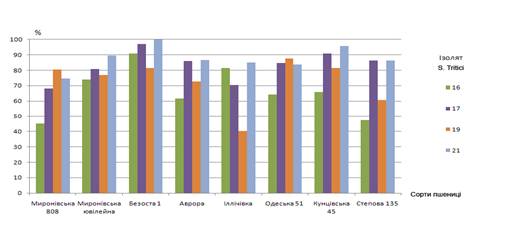
Рис.1. Поразка
листової поверхні сортів пшениці ізолятами Septoria tritici
Трохи більшу
стійкість як по ступеню ураження листової поверхні, так і по інтенсивності
споруляції гриба на рослині показували озимі сорти пшениці в порівнянні з
яровими.
Далі вивчалась
патогенетична дія виділених ізолятів Septoria nodorum на озимі та ярові зернові
культури пшениці. Всі сорти, що брали участь у досліді, були високочутливими до
S.nodorum і уражувалися в значній мірі чотирма ізолятами. Також спостерігалася
кореляція між вірулентністю й інтенсивністю споруляції гриба in vitro. Так,
ізолят 4, що вражав у значній мірі майже всі сорти, що випробовувалися, формував
на них і велику кількість пікнід - до 280 на одному листі, а найменший ступінь
поразки був зареєстрований для ізолята 5 - 46 пікнід на лист (рис.2).
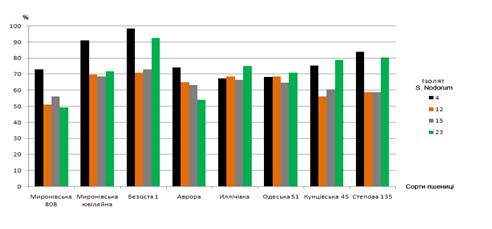
Рис.2. Поразка
листової поверхні сортів пшениці ізолятами Septoria nodorum
Вельми патогенними були ізоляти 4, 12,15 й
23, які в високому ступені (на 51-98 %) уразили 11-12 з 12 досліджуваних сортів
пшениці. Меншу патогенність проявляли ізоляти 1, 5, 14 й 8, що уразили більш
ніж на 50 % лише 6-7 сортів із числа зазначених. Нарешті, найнижчу патогенність
показав ізолят 22, що у сильному ступені уразив лише 3 сорти. Стійкими до
досліджуваних ізолятів Septoria nodorum були Миронівська 808, Аврора,
Кунцівська 45 і Степова 135 за показниками споруляції і цитопатичної дії. Для
окремих ізолятів (наприклад, 12 й 22) відзначений прямий зв'язок між
спорулюючою активністю й патогенністю. Таким чином одержані результати свідчать
про корелятивну залежність ступеню прояву патогенності від біологічних
властивостей збудників септоріозу зернових культур. Створення стійких сортів та
обґрунтування районування їх не можливе без знань складу популяції патогена в
тій чи іншій зоні та систематичного контролю за його змінами. Отже, для
створення ефективних засобів захисту рослин та вивчення стійкості сортів
сільськогосподарських культур до збудників хвороб необхідна розробка точних та
адекватних методів діагностики та визначення морфолого-культуральних ознак
фітопатогенних грибів.
Таким чином,
було встановлено позитивну кореляцію між інтенсивністю спороносіння грибів
Septoria tritici і Septoria nodorum in vitro та проявом їх патогенності.
Показана можливість застосування кількісних показників утворення як пікнід, так
і пікноспор для визначення інтенсивності споруляції та залежної від неї
патогенності. Найвищі показники у досліджуваних ізолятів були зареєстровані на
середовищах – пептонно-сольовий агар та крохмально- глюкозний агар. Виявлено,
що більшому ступеню поразки септоріозом підлягали сорти яриці в порівнянні з
озимими. Відносно стійкими до досліджуваних ізолятів Septoria tritici були
озимий сорт Одеська 51 й яровий Харківська 46, а стійкими до досліджуваних
ізолятів Septoria nodorum були Миронівська 808, Аврора, Кунцівська 45 і Степова
135 за показниками споруляції і цитопатичної дії.