РЕКОНСТРУКЦИЯ
ФИТОЦЕНОЗОВ МАКРОВОДОРОСЛЕЙ В МОРСКИХ ПРИБРЕЖНЫХ ЭКОСИСТЕМАХ
Капков В.И., Сабурин
М.Ю.*, Беленикина О.А., Гусев Д.В.
Московский
государственный университет им. М.В. Ломоносова,
*Всероссийский
научно-исследовательский институт рыбного хозяйства и океанографии
г. Москва,
Россия
В последние годы в
результате возросшей антропогенной нагрузки на морские экосистемы
наблюдается деградация фитоценозов макроводорослей в сублиторали внутренних и
окраинных морей. В этой связи наряду с диагностическими и прогностическими
составляющими биологического мониторинга назрела необходимость реконструкции и
восстановления нарушенных бентосных
фитоценозов, которые оказались наиболее уязвимыми в условиях эвтрофирования и
загрязнения неритической зоны моря.
Макроводоросли бентосных
фитоценозов являются важным компонентом прибрежных морских экосистем. Они
участвуют в трансформации солненчной энергии, продуцируют органическое вещество
и кислород; поглощают из окружающей среды биогенные элементы, ксенобиотики и
участвуют в процессах самоочищения вод; служат субстратом для эпибионтов и
кладок икры беспозвоночных и рыб; являются убежищем для многих гидробионтов.
Многие из них представляют собой ценные промысловые объекты, широко
используемые в различных отрослях промышленности и сельского хозяйства. Однако,
существующие способы добычи промысловых
водорослей и загрязнение морских вод, наряду с другими факторами, способствуют
заметной деградации фитоценозов сублиторали, которая
сопровождается сменой видов-эдификаторов, видового состава, включая
изменение соотношения в сообществе
красных, бурых и зеленых водорослей.
В связи с вышеизложенным
изучение структуры и роли отдельных
видов водорослей сообщества как функциональных доминантов бентосных фитоценозов представляет актуальную
задачу, решение которой будет способствовать пониманию основных причин их
деградации в сублиторали. Такая информация позволит обосновать современные
методы поиска водорослей - биомониторов с неспецифическими откликам на
антропогенные факторы и способы
сооружения и формирования биотической составляющей структуры
искусственных рифов как биологических фильтров в прибрежной зоне моря ( Капков
и др., 2005).
Изучение бентосных
фитоценозов водорослей в сублиторали
Черного моря проводили на специально выбранных полигонах с
использованием легководолазного снаряжения. При визуальном описании фитоценозов
учитывали характер грунта, течения и прибойность, изобатическое распределение
водорослей, их численность и биомассу, проективное покрытие и структуру
сообщества. В работе применяли методику гидроботанических разрезов со
стандартными глубинами от уреза воды до границы произрастания водорослей.
Качественное и количественное распределение фитобентоса в климаксовых
сообществах исследовали в разные сезоны по следующим основным показателям:
видовому и размерно-возрастному составу
водорослей-эдификаторов, водорослей нижних ярусов и эпифитов. Определяли и
рассчитывали площади функционально активных поверхностей таллома
макроводорослей, входящих в фитоценоз. Эксперименты по изучению роста
водорослей и скорости зарастания ими твердых субстратов, а также характер
регенерации талломов после частичного срезания или полного удаления растений
проводили в опытах in situ на глубинах 1-10 м на специальных площадках и
исусственных рифах (Блинова и др., 1990; Сабурин, 2004).
Установлено, что описываемые
ранее двух-, трех- и многоярусные
бентосные фитоценозы макроводорослей в реальности представляют собой чрезвычайно сложные
сообщества с большой активной
поверхностью за счет микро- и
макроэпифитов. При этом площади талломов популяций водорослей претерпевают
существенные изменения в разные сезоны и в зависимости от глу-бины произрастания вида-эдификатора. Максимальные
величины поверхности слоевищ характерны
для конца лета, когда биомасса сезонных летних и одногодичных водорослей
достигала предельных величин (Табл.1).
Таблица 1
Биомасса (г/м²) массовых видов водорослей в
фитоценозе цистозиры
в районе Геленджика
|
Водоросли |
Глубина, м |
|||
|
1 |
3 |
5 |
9 |
|
|
Gelidium latifolium Corallina mediterranea Phyllophora nervosa Ceramium rubrum Apoglossum ruscifolium Polysiphonia subulifera Laurencia coronopus |
81,6±6,8 26,0±2,5 -* 250,5±17,4 -* 5,1±1,2 83,4±6,3 |
310,5±16,0 36,8±3,2 60,1±4,1 28,6±4,4 -* 78,6±4,0 15,6±2,4 |
62,4±4,8 95,7±2,4 90,8±8,3 -* 6,3±1,4 101,7±8,6 -* |
40,6±3,3 53,6±0,7 250,3±18,0 -* 11,2±0,8 12,6±0,8 -* |
|
Сумма биомасс красных водорослей |
446,6±32,2 |
530,3±34,1 |
356,9±25,5 |
368,3±20,3 |
|
Биомасса Cystoseira crinita |
5044,0±184 |
2540,0±82,3 |
605,6±35,6 |
174,2±18,9 |
-* не обнаружено
Так, в фитоценозах сублиторали Черного моря на твердых субстратах по
видовому обилию, а с глубиной по
биомассе и поверхности талломов преобладали красные водоросли с рассеченными
слоевищами, выполняющие роль функциональных доминантов в сообществе (Рис.).
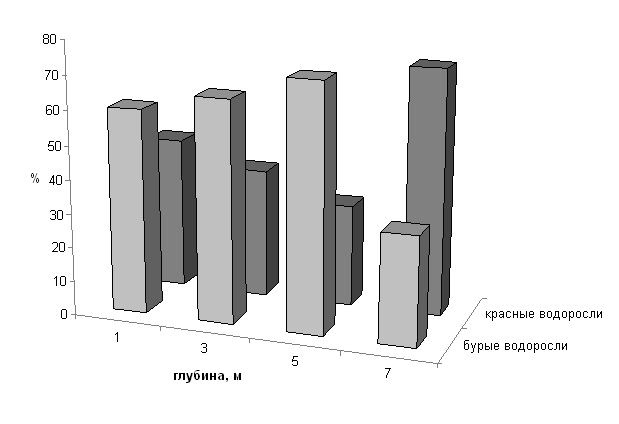
Рис. Поверхности талломов популяций красных и
бурых водорослей в фитоценозе цистозиры
Функционально активные поверхности макроводорослей
значительно увеличивались за счет эпифитных форм диатомей (Табл.2). Обычно на
красных, бурых и зеленых макроводорослях с шероховатыми талломами обитали
многочисленные одноклеточные (Navicula directa, Licmophora ehrenbergii) и колониальные
(Synedra tabulata, Achnanthes brevipes) бентосные
диатомовые водоросли. На гладких покрытых слизью талломах поселялась плоская по форме одноклеточная
диатомея Cocconeis scutellum (Беленикина, 2005).
Таблица
2
Изменение
биомассы и поверхности талломов цистозиры и красных
водорослей, входящих в фитоценоз на разных глубинах в районе мыса Б.Утриш.
|
Глубина, м |
Cystoseira crinita |
Водоросли нижних ярусов |
Водоросли-эпифиты |
|||
|
Биомасса, г/м² |
S таллома, м² |
Биомасса, г/м² |
S таллома, м² |
Биомасса, г/м² |
S таллома, м² |
|
|
1 3 5 7 9 |
6920±84 8163±88 8609±53 2845±41 1250±25 |
65,7 77,5 81,7 27,0 11,8 |
754±15 609±12 910±16 816±18 681±14 |
7,5 35,2 12,8 9,1 8,3 |
311±12 2032±29 3109±33 4211±48 1552±24 |
31,8 56,7 77,3 81,5 64,6 |
При исследование динамики восстановления нарушенных
фитоценозов бурых водорослей обнаружена
их высокая способность к регенерации. На экспериментальнвх площадках в
море на глубинах 0,5 - 7 м рост стволиков, боковых и адвентивных ветвей у бурой
водоросли Cystoseira crinita после срезания на высоте 1-5 см от подошвы
наблюдался спустя 2-3 месяца. К этому времени на свободных участках субстрата
появились проростки цистозиры длиной 25 мм, которые развились из зигот.
Восстановлению зарослей цистозиры способствовали также регенерирующие из
базальных пластин отростки слоевищ. К
концу первого года от начала эксперимента длина стволиков достигала почти 100
мм, на которых насчитывалось до 10-15 боковых ветвей. Через два года длина стволиков достигала 150 мм, а боковых и
адвентивных ветвей 300 и 30 соответственно. На отдельных боковых ветвях обнаруживались рецептакулы.
Водоросли на экспериментальном участке практически не отличались от соседних талломов климаксового фитоценоза
цистозиры.
После полного
удаления водорослей с субстрата восстановление растительного сообщества
происходило исключительно за счет прорастания
зигот. После выхода яйцеклетки из концептакул и оплодотворения, зигота
покрывалась плотной коричневой оболочкой и после оседания на субстрат выделяла
слизь и становилась неподвижной. Через неделю зигота превращалась в проросток,
прикрепляясь к субстрату мощными
ризоидами. У Cystoseira crinita, проирастающей
в основном в зоне морского прибоя ризоиды
были толщиной 50-60 мкм, с грушевидными вздутиями, перегородками и боковыми
выростами что увеличивало площадь контакта проростка и слоевища с субстратом.
Спустя два года рост талломов происходил с той же скоростью, что и на соседних участках у водорослей зрелого
сообщества. При этом шло формирование многоярусного хотя и несколько
обедненного по видовому обилию
фитоценоза с измененной ярусностью и биомассой. На цистозире летом
поселялись многочисленные
водоросли-эпифиты, среди которых преобладали багрянки: Ceramium rubrum, Polysiphonia sanguinea, Laurencia coronopus и другие ( Табл.3).
Таблица 3
Восстановление фитоценоза Cystoseira crinita
после полного удаления растительности с
субстрата
|
Время роста, мес. |
Биомасса, г/м² |
Плотность, экз/м² |
Длина цистозиры, см |
||
|
C .crinita |
Водоросли нижнего яруса |
стволик |
таллом |
||
|
3 9 12 24 |
692 5630 5250 5867 |
28 530 950 217 |
480 3600 3400 2800 |
1,7 2,6 5,4 12,5 |
10,0 10,8 14,9 17,5 |
В сублиторальной зоне моря отсутствие необходимого твердого субстрата часто
оказывается основным фактором, ограничивающим развитие донных фитоценозов.
Разорванность бентосных растительных сообществ обусловлена именно мозаичностью
распространения каменистых субстратов, которые предпочитают большинство бурых и
красных водорослей. Сооружение искусственных рифов (ИР) позволяет увеличить
площадь субстрата, на котором растут
макроводоросли. В результате проведенных
исследований было обнаружено, что формирование фитоценозов бурых и красных
водорослей на ИР на глубинах 1-10 метров составляло около трех лет. В Черном
море видами эдификаторами таких фитоценозов чаще всего были Cystoseira crinita и Phyllophora nervosa с общим числом
видов в сообществе до 80% и близкими
величинами биомассы. При этом была установлена последовательность заселения
каменистого субстрата ИР, которая в общих чертах повторяла колонизацию
водорослями очищенных от растительности естественных твердых субстратов.
Первыми поверхности ИР занимали бентосные одноклеточные подвижные формы
диатомей Navicula pennata и
N. directa. Затем в
обрастаниях появлялись неподвижные Cocconeis scutellum и
Hyalodiscus ambiguus, а также
колониальные формы диатомовых водорослей Achnanthes brevipes , Synedra tabulata и Licmophora sp. На покрытом
диатомеями субстрате селились корковые и стелющиеся формы багрянок Lithothamnion lenormandi, Phymatolithon polymorphum, Rhodochorton penicilliforme, которые
формировали нижний ярус сообщества. Формирование последующих ярусов фитоценоза
происходило практически одновременно. Верхний
ярус, создающий основной фон сообщества, создавали цистозира или
филлофора, под покровом которых развивались водоросли второго и третьего ярусов
красные Gelidium latifolium, Corallina mediterranea, Ceramium rubrum, Laurencia obtusa и бурая водоросль Cladostephus verticillatus в фитоценозе филлофоры. Эпифитные синузии на
макроводорослях всех ярусов появлялись весной, когда начинался интенсивный рост сезонно-летних и
одногодичных водорослей. Практически во всех фитоценозах, которые
сформировались на ИР, красные водоросли преобладали по видовому обилию и
величине активной поверхности талломов. Так, в фитоценозах цистозиры биомасса
бурых водорослей была в 3 раза больше, чем красных, в то время как площадь
талломов последних превышала 70% общей поверхности слоевищ фитоценоза. Кроме
того, талломы макроводорослей обрастали эпибионтами, включая моллюсков-фильтраторов. В результате, на
искусственных рифах формировались сложные биоценозы, макроводорослей с рассеченными талломами, на
которых поселялись микро- и макроэпифиты, а также многочисленные
моллюски-фильтраторы, выполняющие в сублиторали функцию мощных биологичнских фильтров ( Капков и др., 2006).
Таким
образом при осуществлении биологического мониторинга сублиторальной зоны моря
при освоении шельфа, сооружение
искусственных рифов с большим числом функциональных доминантов –
красных, бурых и зеленых макроводорослей с высокой удельной поверхностью
талломов, включая многочисленных эпибионтов, способствует, как показали наши
исследования, изъятию из окружающей
среды органических и
неорганических веществ, включая тяжелые металлы-ксенобиотики, что ведет к
снижению уровня эвтрофирования и
загрязнения прибрежных морских экосистем.
Литература.
Беленикина О.А. Красные водоросли в системе биомониторинга сублиторали
Черного моря. Автореф. дисс. канд. биол. наук. М. 2005. 20 с.
Блинова
Е.И., Пупышев В.А., Сабурин М.Ю., Тришина (Беленикина) О.А. Фитобентос искусственных рифов
прибрежных вод черноморского побережья северного Кавказа. Сб. научн. труд..
ВНИРО «Искусственные рифы для рыбного
хозяйства». М., 1990. С. 127-141.
Капков
В.И., Сабурин М.Ю., Беленикина О.А., Блинова Е.И. Восстановление фитоценозов Cystoseira crinita (Phaeophyta) и динамика роста макрофитов на
искусственных рифах. Вестн. Моск. ун-та. Сер. 16. Биология. 2005. № 2. С.
30-34.
Капков
В.И., Беленикина О.А., Сабурин М.Ю. Искусственные рифы в биомониторинге прибрежных морских
экосистем. Сб. научн. труд. междунар. конф. «Современные проблемы адаптации и
биоразнообразия». Махачкала, 2006. С.17-19.
Сабурин
М.Ю. Фитоценозы
черноморской цистозиры: структура, восстановление и перспективы использования.
Автореф. дисс. канд. биол. наук. М., 2004. 24 с.