Ph.D. in Biology, Murylev A. V.
Perm State Humanitarian
Pedagogical University, Russia
SUPERCOOLING
POINTS OF APIS MELLIFERA MELLIFERA
AND APIS MELLIFERA CARPATHICA
INTRODUCTION
Perm krai marks
the northern limit of the natural distribution range of honeybees. Honeybees of
the Apis mellifera mellifera L.
successfully overwinter in this area and display high degrees of productivity.
In the Perm krai Region, the Kama Region population of A. m. mellifera has been recognized [7], distinguished by
morphological and ecophysiological characters from other A. m. mellifera populations
of Russia. Honeybees of this population have a genetically secured complex of
adaptations to northern environmental conditions. In addition to the Perm krai
Region population of A. m. mellifera,
some beekeepers have started breeding at their apiaries the more peaceful Apis mellifera carpathica. The A. m. carpathica have been brought to
the Perm krai Region for this experiment from the Mukachevo Purebreed Bee
Nursery, Ukraine. It is known that not all organisms are able to successfully
adapt to new ecological and geographical conditions, however, some positive
qualities encourage beekeepers to acclimatize separate races of bees outside
the borders of their natural habitats. The adaptations of these bees evolved
under the conditions of their southern range and are distinguished by some
peculiar features. They have a body size smaller than that of the A. m. mellifera, but their proboscis
length is slightly longer, allowing them to use a wider range of meliferous
plants [2]. Ecological and biological characteristics of bees Carpathian race
for them in unusual climatic conditions remain poorly studied.
The cold-hardiness
of insects is usually estimated by the supercooling point (SCP), a parameter of
proposed by Bakhmet'ev [1]. Crystallization is usually lethal to the insect,
because it damages the tissues of its body. In the light of new technical
abilities and the emergence of new technologies, the study of the SCP parameter
continues also at the current stage of scientific development [4; 5]. It is
especially important for characterizing different honeybee subspecies [6].
The purposes of
this study were to analyze the resistance to low temperatures in honeybees of
the Kama Region population of the A. m.
mellifera and those of the A. m.
carpathica on individuals of the spring, summer, and autumn generations and
the total percentage of water in the body of honeybees during different periods
of bee family development.
MATERIALS AND
METHODS
The experiments
were performed monthly from 2010 to 2013. The materials included bees of the
Kama Region population of the A. m.
mellifera from the Nizhnesypovskoe Bee Breeding Farm (Uinskoe district,
Perm krai) and bees of the A. m.
carpathica brought to Perm krai from the Mukachevo Purebreed Bee Nursery,
Ukraine.
Instruments and testing methods for the testing of SCP
SCP was measured
according to the method proposed by Es'kov [3], in each of the three tagmata
(head, thorax, and abdomen). A total of 20 bees of the studied races were used
every month. The measurements were taken using a chrome/copel thermocouple
fixed on a wooden base. The thermocouple was attached to one of the body parts
of bees vaseline, and then placed in the freezer compartment of the refrigerator.
The temperature values of SCP were recorded with a Termodat-38M2 device. SCP is
determined by measuring the heat released in the course of crystallization,
when matter transforms from the liquid to the solid state (fig. 1). Simultaneously,
SCP was compared to the total water content in the body of honeybees of
different generations.
Testing of water content
The total amount
of water in the body was determined as the difference between two results of
weighing the bee's body: wet weight and constant weight after drying the
specimen at 102°C for 48 hours.
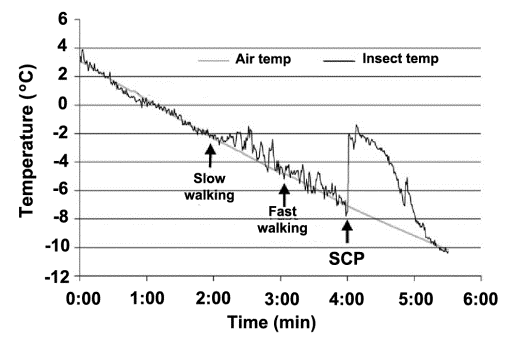
Fig. 1. Supercooling
point of insects
RESULTS
Research results
showed that the total water content in the body of honeybees changes over the
year. With the cessation of flying activity in September or October and gradual
advent of the cold period, the water content in the body of bees of the autumn
generation decreases by 8 %, leading to a slowdown of metabolic processes. Bee
families enter a period of physiological dormancy of development and reproduction.
During this period, a decrease in the supercooling point has been recorded
(table).
In February, bees
display the lowest water content in the body; by this time, the supercooling
point also reaches its lowest subzero values. Significant differences in these
parameters have been found between the studied races in bees of the autumn
generation (t = 4.08, p<0.01). In
April, overwintered bees are replaced by young bees of the spring generation.
The young bees
display a higher water content in the body and an increase of SCP in the
tagmata. During the period of intense growth and accumulation of inactive bees,
which start working during the period of principal honey flow, the supercooling
point in the tagmata continues growing. Bees of the summer generation emerge,
which display the highest water content in the body and the highest values of
SCP. These parameters remain high until October, and then decrease again. In
bees of the summer generation, differences between the subspecies proved
insignificant (t = 2.48, p>0.05). Positive correlation has been observed
between the values of SCP and the total water content in the body: the
correlation coefficient is 0.84 in the Kama Region bees of the A. m. mellifera and 0.92 in those of the A. m. carpathica.
Dynamics of SCP and water content in the body of bees
of the Apis m.
mellifera and Apis m. carpathica
|
Generation of bees |
Apis
m. mellifera |
Apis
m. carpathica |
||||||
|
Water content, %, n = 180 |
SCP, (Т±m), °С, n =
180 |
Water content, % |
SCP, (Т±m), °С, n = 180 |
|||||
|
head |
thorax |
abdomen |
head |
thorax |
abdomen |
|||
|
Spring |
65.67 |
–7.45± 0.08 |
–5.67± 0.10 |
–4.93± 0.02 |
71.29 |
–7.01± 0.12 |
–5.56± 0.12 |
–5.01± 0.04 |
|
Summer |
72.42 |
–4.45± 0.02 |
–4.56± 0.07 |
–3.98± 0.08 |
74.19 |
–4.42± 0.04 |
–4.50± 0.01 |
–4.02± 0.04 |
|
Autumn (October-December) |
64.31 |
–7.83± 0.12 |
–6.68± 0.09 |
–5.81± 0.09 |
69.25 |
–7.12± 0.13 |
–5.81± 0.08 |
–5.54± 0.11 |
|
Autumn (January-March) |
63.22 |
–10.61± 0.06 |
–8.25± 0.05 |
–8.04± 0.07 |
67.34 |
–9.74± 0.09 |
–7.12± 0.12 |
–7.24± 0.10 |
Analysis of SCP
values recorded in different tagmata of honeybeesshows that in the Kama Region
population of the A. m. mellifera the
lowest values of SCP in the head were observed in February, reaching -10.61 ±
0.06°C (n = 180), while in the A. m.
carpathica the lowest values were observed in March, reaching -9.74 ± 0.09°C
(n = 180) (table). The values of SCP in the thorax were higher: in bees of the
Kama Region population of the A. m.
mellifera they reached ‑8.25 ± 0.05°C (n = 180) in February, and in
those of the A. m. carpathica they
reached -7.12 ± 0.12°C (n = 180) in March. In the abdomen, the lowest values of
SCP in the Kama Region population of the A.
m. mellifera were observed in March, reaching -8.04 ± 0.07°C (n = 180); in
bees of the A. m. carpathica they
were also observed in March, reaching -7.24 ± 0.10°C (n = 180). In addition, it
has been noted that at the end of overwintering A. m. carpathica switch to the active state 15 to 20 days earlier
than A. m. mellifera ones. Bees of
the former race start nurturing their brood already in March, according to
their genetic program, although the flying period starts only some 20 days
later. A. m. mellifera start
nurturing their brood only after the start of their flight. In March they continue
to build a winter club.
CONCLUSIONS
The lowest
supercooling point values have been recorded in bees with the lowest water
content in the body. The water content in the body of bees from the Kama Region
population of the A. m. mellifera is
lower during the entire annual cycle than in those of the A. m. carpathica. Summer bees had the highest SCP temperature and
high water content. It has been shown that A.
m. carpathica are less physiologically adapted to long overwintering under
low temperatures. In combination with other factors (such as diseases or the
duration of the flightless period), these parameters can have an adverse effect
on the overwintering of A. m. carpathica
in Perm krai.
REFERENCES
1. Bakhmetyev P.
(1898) – The temperature of insects. Scientific Review., 5: 1602–1611.
2. Bodnarchuk L.
I., Gaidar V. A., Pilipenko V. P. (2008) – Carpathian bee, what are they? Bee J., 2: 1–2.
3. Eskov E. K. (1991)
– Methods and techniques of experimental zoology. Ryazan. ped. Inst. Press, Ryazan.
4. Eskov E. K.
(2007) – The temperature of maximum supercooling and the state of the fat body
of bees. J. Beekeeping., 6: 22–23.
5. Heinrich B.
(1993) – The Hot-blooded Insects, Strategies and Mechanisms of
Thermoregulation, Springer Press, Heidelberg, Berlin.
6. Murylev A. V.,
Petukhov A. V. (2011) – The point of crystallization of different parts of the
body bee populations Kama. J. Beekeeping,
1: 18–19.
7. Shurakov A. I.,
Petukhov A. V., Eskov E. K. (1999) – Saving the gene pool of the Central
Russian bees, and basic directions of development of beekeeping in the Perm
region. Perm. State. ped. University Press, Perm.