Д.б.н.
Воробьёва Т.М., к.б.н. Бевзюк Д.А, Левичева Н.А.
ГУ “ Институт
неврологии, психиатрии и наркологии НАМНУ”, Украина
Биоинформационный
подход к купированию агрессивного поведения у крыс.
В современном социуме одной из важнейших проблем бытия
является галопирующий рост агрессии. Системообразующим фактором последней
является модель будущей положительной эмоции в акцепторе результата действия.
Это приводит к криминалу, садизму, насилию, аутоагрессии и другим
антисоциальным действиям [2, 4]. Поэтому необходимы исследования углубленных
механизмов формирования агрессивности (эмоционально негативной реакции) и
агрессивного поведения (мотивации). В этой связи приобретают значения вопросы
использования новых нанотехнологий, содержащих биологически информационное
квантирование: путем запуска атомно - молекулярной конструкции генетических
кодонов памяти, - носителей частной программы эволюции, заданной, приобретенной,
модулированной пространственным энергоинформационным гомеостазом. В
экспериментальных исследованиях показана возможность переноса информации
заключённой в биопотенциалах мозга. С помощью технических магнитных или других
носителей она может храниться и передаваться дистанционно реципиенту от донора
и управлять поведением животного организма [1].
Материалы и методы.
Эксперимент был проведен на 28-ми крысах -
самцах 5 -ти - 6-ти месячного возраста. Животным осуществляли стереотаксическую
имплантацию нихромовых электродов в лимбико - неокортикальные структуры мозга
билатерально: лобно - фронтальный неокортекс, гиппокамп, гипоталамус и septum
под кетаминовым наркозом. Электрическая стимуляция заднего вентромедиального
гипоталамуса (ЗВМГПТ) приводила к появлению пароксизмальных отрицательных
реакций, а её электрографическими коррелятами являлась пароксизмальная и
судорожная активность на ЭЭГ. Самостимуляцию положительных эмоциогенных
структур мозга осуществляли в камере Скинера по методу[4] .Затем осуществляли
запись ЭЭГ с введением информации в блок компьютерного электроэнцефалографа («запоминающего
устройства»), где она может храниться в «банке». С этого «банка» информация, заключенная
в биопотенциалах мозга может извлекаться и использоваться в определённое время
для коррекции модельных форм поведения, в конкретном случае агрессивного
поведения.
Регистрацию ЭЭГ проводили с помощью компьютерного
электроэнцефалографа «Нейрон -спектр+». Анализ 5 фрагментов ЭЭГ эпохами по 2 секунды осуществляли визуально и с помощью
математического анализа с применением пакета компьютерных программ. Статистическую
обработку данных осуществляли с помощью программы Statistica 6.1 с
использованием непараметрического критерия Вилкоксона.
Результаты исследований
1 2 3 4 А В Б
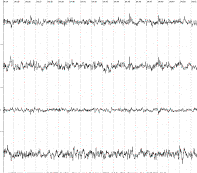
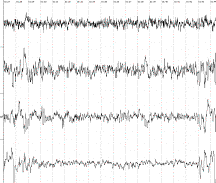
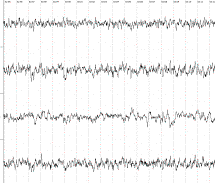
В фоновой ЭЭГ исследуемых крыс
в неокортексе доминировала смешанная активность с эпизодическими комплексами ,
включающие тета- , дельта -волны, острые разряды.
1-лобно-фронтальный неокортекс, 2-гиппокамп, 3-гипоталамус,
4–septum.
А-фоновая ЭЭГ, Б – после стимуляции
вентромедиального гипоталамуса, В- после введения информации заключённой в
биопотенциалах ЗВЛГПТ крысы донора.
Рисунок 1.
Динамика изменений электрической
активности мозга на примере крысы № 16 после воздействий.
В гиппокампе в подавляющем большинстве случаев
доминировала низкоамплитудная альфа активность. В гипоталамусе на фоне
смешанной активности доминировала низкоамплитудная и высокочастотная активность
в диапазоне альфа-бета-волн.
В septum в 90%
случаев на фоне смешанной активности участки бета-активности постоянно
перемежались с участками тета- и дельта- волн, формируя комплексы .
Принудительная стимуляция
вентромедиального гипоталамуса у исследуемых крыс вызавала появление негативных
эмоциональные реакции, зачастую переходящих в пароксизмы агрессии. При этом в
электрической активности мозга испытуемых животных происходили изменения. В неокортексе
в 70 % случаев преобладала низкоамплитудная высокочастотная активность в
диапазоне бета. В гиппокампе в 80 % крыс регистрировались комплексы
медленноволновой активности на фоне смешанной активности. В гипоталамусе у 80%
крыс на фоне резко сниженного электрогенеза отмечались единичные пик - тета-
(рис. 1, Б ). В septum у всех крыс регистрировалась медленноволновая
активность,чередующаяся с небольшими эпохами бета - ритма и комплексами сложной
структуры (рис. 1, Б ).
Спектральная мощность колебаний
тета- , альфа - и бета диапазона при стимуляции вентромедиального гипоталамуса
показала достоверный рост этого показателя по сравнению с фоновыми.
После проведенного курса лечения
при котором в реальном режиме времени крысам реципиентам вводилась информация,
находившаяся в биопотенциалах ЗВЛГПТ крысы –донора выявлены изменения в
электрогенезе лимбико-неокортикальных структур мозга и поведении исследуемых
животных. Выбор ЗВЛГПТ был обусловлен тем, что он является триггерной
структурой в формировании сложной морфофункциональной системной организации
положительных эмоций .
На электроэнцефалограмме
исследуемых крыс (пример ЭЭГ крысы №16) после введения ЭЭГ донора было
зарегистрировано снижение амплитуды альфа - и тета – активности. Исчезли
комплексы, значительно уменьшилось количество острых волн. ( рис. 1 В),было
выявлено снижение спектральной мощности тета- ритма в гиппокампе , а также
альфа - тета- ритмов в septum в сравнении со стимуляцией ЗВМГПТ. Животные
становились спокойными , длительное время сидели неподвижно , регистрировался
положительный груминг. В заключении следует сказать, что принудительную
стимуляцию ЗВМГПТ можно использовать как адекватную модель отрицательного
эмоционального состояния, про это свидетельствует прямая корреляция
поведенческих показателей (реакций избегания, усиления вокализаций,
отрицательного груминга) пароксизмальных аффектов эмоциональных негативных
реакций с показателями электрической активности лимбико-неокортикальных
структур моза. А введение информации из положительнай эмоциогенной зоны ЗВЛГПТ
от крысы донора крысам реципиентам уравновешивает эмоциональный гомеостаз, что
выражается в исчезновении судорожных компонентов и диффузных аффективных
пароксизмов на ЭЭГ, проявлениях положительного груминга и спокойствия. Этот
способ купирования отрицательного эмоционального агрессивного поведения
представляется нам эффективным для дальнейшего изучения, как нового способа
коррекции различных эмоциональных состояний.
Литература
1. БакуменкоЛ.П., Верзин И.И., Воробьева Т.М., Дабагян
А.В., Кононенко И.В., Лещенко А.Г. Авторское свидетельство № 908378, 1981.
Способ управления поведением животного организма. БакуменкоЛ.П., Верзин И.И.,
Воробьева Т.М., Дабагян А.В., Кононенко И.В., Лещенко А.Г., Напеч. 02.11.1981
2. Бевзюк, Д.А., 2012. Нейрофизиологические
механизмы трансформации социофобий в агрессивное поведение в условиях
депривации гедонического поведения. Український вісник психоневрології, т. 20, вип. 3, 162
3. Воробьева, Т.М., Бевзюк Д.А, Кириллова Н.А., 2009.
Роль эмоциогенных стктур мозга в механизмах формирования фобий и их возможной
трансформации в агрессивное поведение у крыс. Психическое здоровье, №4, 46-51.