Биологические
науки/5.Молекулярная биология
Bulgakova O.V., Bersimbay R.I.
mTOR Complex 2 Is Required for the Development
of Epithelial cells in Mice.
L.N. Gumilyov
Eurasian National University, Kazakhstan
mTOR belongs to the family of phosphoinositide-3-kinase
(PI3K)-related kinases (PIKKs), along with ATM, ATR, DNA-PK, and hSMG1[1]. All
of these proteins have C-terminal protein kinase domains with similarity to the
lipid kinase PI3K, thus giving the family its name mTOR forms two different
protein complexes defined by the proteins to which it is bound, exerting
different but related functions [2]. The mTOR complex 1 (mTORC1) is defined by
the presence of Raptor, mLST8/GβL, Deptor and
Pras40 [3], whereas Rictor, GβL, Protor,
Deptor and mSin form mTORC2 [4,5]. mTORC1 regulates protein biosynthesis through phosphorylation of the ribsosomal
protein S6 kinase (p70S6K) and subsequent phosphorylation of the ribosomal
protein S6 (rpS6), or
phosphorylation and inactivation of the eukaryotic initiation factor 4E
(eIF4E)-binding protein-1(4E-BP1), dissociating 4E-BP1 from the RNA cap-binding protein eIF4E, thus promoting
cap-dependent translation of mRNAs [6]. In contrast to mTORC1 complex, mTORC2 is
currently poorly understood. Rictor
is the important component of
mTORC2 [4].To
determine the role of mTORC1 in development, we studied the effects of deleting Rictor on
embryogenesis.
Results
The Cre/loxP system was used to develop conditional Rictor KO
mutants. A previously established mutant mouse line was used, n which Rictor
was flanked by 2 loxP sequences. To
obtain conditional Rictor KO, we then
crossed mutant mice harboring the Rictor floxed allele with mice, which have Cre- recombination. to deleted of.
When expressing Cre recombinase started under the control of the K14
promoter the loxP-flanking region was deleted. Genotypes were confirmed by PCR
of tail DNA, using oligonucleotide primers pairs for floxed Rictor and for the Cre
transgene. The genotyping was confirmed by Western blot of the total lysates of
the embryos epithelial cells to show
that the Akt/PKB expression.
In
these studies, we generated mice deficient for rictor in epithelial tissues to determine
the role of mTORC2 in the embryonic development of mammals.
Immunoblot analyses of protein lysates prepared from rictor-deficient
epithelial cells reveal that gene products are necessary for the basal
phosphorylation of Akt/PKB on S473 of its hydrophobic motif. Rictor-deficient embryos die at early
stages of development (8.5 days).
We conclude that rictor is an
essential component of mTORC2 and is required in early development.
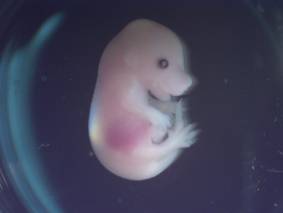
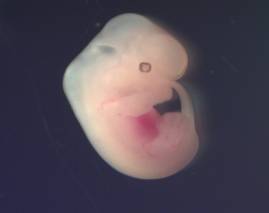
A B
Fig. 1 Phenotypic
Characterization of Embryos (A) – 15.5 days, (B) – 8.5 days
REFERENCES:
1.
Harris T.E.,
Lawrence J. TOR signaling.// Sci.STKE 2003. Vol.2003 , P.212-215
2. Kim D.H., Sarbassov D.D., Ali S.M., King J.E., Latek R.R.,
et al. mTOR interacts with raptort of ormanutrient-sensitive complex that
signals to the cell growth machinery.//Cell. 2002. Vol. 110. P.163–175
3.
Gingras A., Baught
B., Sonenberg N. Regulation of translation initiation by FRAP/mTOR.//Genes
&Development. 2001. Vol. 15. P. 807-826
4.
Sarbassov D.D.,
Ali S.M., Kim D.H., Guertin D.A., Latek R.R., Erdjument-Bromage H., Tempst P.,
and Sabatini D.M. Rictor, a novel binding partner of mTOR, defines a
rapamycin-insensitive and raptor-independent pathway that regulates the
cytoskeleton.// Curr.Biol. 2004. Vol. 14. P. 1296–1302
5.
Sarbassov D. D.,
Guertin D. A., Ali S. M. and Sabatini D. M. (2005).Phosphorylation and
regulation of Akt/PKB by the rictor-mTOR complex. //Science. 2005. Vol. 307. P.
1098-1101
6.
Gingras A., Baught
B., Sonenberg N. Regulation of translation initiation by FRAP/mTOR.//Genes
&Development. 2001. Vol. 15. P. 807-826