Давидчук
Н.В., Медведев Д. А.
Тамбовский
государственный университет им. Г.Р.Державина
О возможности изменения некоторых
физико-химических показателей воды
водными растениями
В естественных условиях физико-химические
показатели воды могут изменить как проплывающие рыбы, выбросы крупного
предприятия, так и водные растения.
Поскольку гидрофиты никогда не
испытывают недостатка в воде, лимитирующим в их развитии становится фактор
питания. Потребляя днем растворенный углекислый газ СО2, растения
снижают содержание его в воде и оказываются в неблагоприятных условиях.
Считается, что при рН>8,3
(точка эквивалентности
фенолфталеина) свободная
углекислота в воде практически отсутствует [1]. Впрочем, это касается мягкой воды, в
которой растворено мало солей кальция. В водах, богатых кальцием, водные
растения могут продолжать потребление, добывая углекислый газ из соединений
кальция, из кислот и других соединений углерода. Очевидно, что добывать
углекислый газ этими путями водным растениям значительно сложнее, чем растениям
суши. И не все погруженные гидрофиты в равной степени способны насыщаться путем
разложения сложных веществ. В водах,
богатых известью, растения, способные усваивать бикарбонатный ион СО2,
выделяют его из группы HCO3- , при этом образуются ионы ОН−,
которые подщелачивают воду во время фотосинтеза до 10 – 10,4 рН [2]. Чем
меньше водоем, тем резче колебания рН в воде ночью и днем. Помимо этого снижается временная
(карбонатная) жесткость воды за счёт уменьшения количества карбонатов в воде и
выпадением СаСОз в осадок
( процесс биогенного умягчения воды) [3]. Биогенное умягчение тем больше, чем выше
освещенность растений в аквариуме. По этому процессу умягчения воды определяли
активность разных видов гидрофитов в использовании бикарбонатов. Выяснилось,
что активность эта бывает разной не только у разных видов, но и у одного вида
при разной освещенности. Растения в домашних условиях из-за низкой
освещенности в большинстве своем после усвоения свободного СО2 явно
недостаточно используют бикарбонаты и, следовательно, довольно ощутимо
голодают. Другим тормозом в
ассимилятивной деятельности гидрофитов является неподвижность воды. Если держать водные растения постоянно в застойной
воде, то явные признаки голодания обнаруживаются даже тогда, когда остальные
факторы (свет, температура, наличие карбонатов и т. д.) оптимальны.
Однако объективно оценить влияние растений
на состав воды довольно сложно. Тем не менее, технология ризофильтрации
позволяет выявить некоторые закономерности изменения состава воды за счёт
деятельности корней водных растений, которые
способны поглощать,
концентрировать и осаждать из водных растворов опасные химические соединения
[4]. Все эти процессы протекают при
определённом значении pH водной среды,
поэтому поддержание данного показателя на оптимальном для растений уровне
весьма важно.
Целью нашего исследования явилось
получение сведений о суточном изменении таких показателей воды как рН и общей жесткости (dGH) водными растениями.
Эксперимент проводился в лабораторных условиях.
Объектами послужили мелколистные водные растения - типичные представители гидрофлоры Центрального Черноземья: роголистник
погруженный – (Ceratophyllum demersum L.), элодея канадская – (Elodea canadensis Michx) [5], а также представитель флоры тропических водоемов Амазонки - перистолистник бразильский – (Myriophyllum aquaticum Cambeesedes).
Определенное количество растений одного
вида погружалось в заполненные водой (pH – 8, dGH – 160) цельностеклянные аквариумы объемом
10 л. Над аквариумом устанавливалась люминесцентная лампа flora-glo
производства фирмы Hagen имеющая спектр 2800К, 90 LUX.
Световой день регулировался при помощи
электронного таймера производства фирмы Quigg. Аквариумы освещались непрерывно в течение 12 часов.
Каждые два часа из всех трех аквариумов производился забор воды. Анализ производился при помощи экспресс
тестов фирм Tetra (рН тест) и НПФ «Aquatest» (dGH тест) в
соответствии с прилагающейся инструкцией по применению. Температура воды при
этом постоянная - 20 0C.
В результате измерения pH было
выявлено, что в течение светового дня значения рН практически не изменялись,
только в самом конце периода освещения начали расти. Рост значений продолжился
и после прекращения освещения, и шел на протяжении 5 часов. Полученные данные
представлены на рисунке 1.
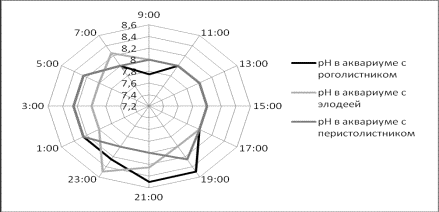
Рисунок 1. Значения pH в пробах воды в аквариумах за сутки.
Значение dGH изменялось в обратной зависимости от рН. А
максимальные значения рН отмечались перед всплесками жесткости после
прекращения светового дня. В дневное время
общая жесткость постоянно падала (рис.2).
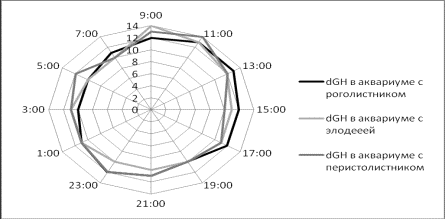
Рисунок 2. Значения dGH в пробах воды в аквариумах за сутки.
В наших исследованиях к концу светового дня наметилась явная
нехватка углекислого газа, но если элодея и роголистник для получения
углекислого газа начали преобразовывать гидрокарбонат кальция, то для
перистолистника такой процесс не характерен, что, и наблюдаем на графике
изменения рН. Полученные данные свидетельствуют о том, что в условиях
лабораторного эксперимента большей адаптационной способностью обладают элодея и
роголистник.
Кроме того, в первом и во
втором аквариуме в результате произошло падение значения dGH, которое после отключения света стало
восстанавливаться, а значение рН постепенно снижаться.
В целом проведённый
эксперимент дает представление об общей закономерности циклических процессов,
происходящих в водных экосистемах.
литература
1.
Яночкин А. О жесткости, рН, СО2 и
буферных системах… //Аквариум.2005. №6. С 40 – 46.
2.
Грошина Т.К. Экология растений. Учебное пособие. М.:
Высшая школа, 1979. 368с.
3.
Махлин М.Д.
По аллеям гидросада М.: 1984.
4.
Прасад М.Н. Практическое
применение растений для восстановления экосистем, загрязненных металлами. //
Физиол. растений. Обзор. 2003, том 50, №5, с.764-780.
5.
печенюк Е.В. Атлас высших водных и пребрежно-водных
растений. Воронежский госпедуниверситет, 2004. 129 с.