Биологические науки / 6. Микробиология
Брюховецька Д.О.1, Броварська О.С.2,
Варбанець Л.Д. 2, Воронкова О.С. 1, Вінніков А.І. 1
Дніпропетровський національний університет ім. Олеся
Гончара, Україна
Інститут мікробіології і вірусології ім. Д.К. Заболотного
НАН України,
м. Київ, Україна
БУДОВА ТА ВЛАСТИВОСТІ ЛІПОПОЛІСАХАРИДУ
БАКТЕРІЙ
Вступ
Взаємодія мікробної клітини із
зовнішнім середовищем зумовлена будовою її поверхневих структур. Грамнегативні
бактерії мають особливу типову будову клітинної стінки, зовнішній шар якої
складається із біополімерів, які представлені у тому числі глікополімерами.
Серед них важливу роль відіграють ліпополісахариди (ЛПС). Ці біополімери
являють собою основний інгредієнт клітинної оболонки грамнегативних бактерій.
Перебуваючи на поверхні мікробної клітини і взаємодіючи з іншими біополімерами,
вони стабілізують зовнішню мембрану, виконуючи конструктивну роль; здійснюють
бар'єрну функцію, перешкоджаючи попаданню в мікробну клітину ядів, детергентів,
антибіотиків та інших лікарняних препаратів; служать акцепторами бактеріоцинів
і фагів тощо.
Являючись при цьому основними
термостабільними антигенами і ендотоксинами мікробної клітини, ЛПС визначають
її серологічну специфічність, О- антигенність і значною мірою - характер
взаємодії з іншими мікроорганізмами або макроорганізмом у біоценозах, беруть
участь в інфекційному процесі в різних біологічних системах [1].
Мета роботи – охарактеризувати
будову та біологічну активність ліпополісахаридів грамнегативних бактерій.
Загальне уявлення про ліпополісахарид
Ліпополісахариди – це
термостабільні біополімери, що виробляються грамнегативними бактеріями. Вони є
основною поверхневою структурою грамнегативних бактерій, яка відіграє важливу
роль у взаємодії мікро- та макроорганізму. Безпосередній контакт молекули із
зовнішнім середовищем значною мірою зумовлює функції цих біополімерів у
мікробній клітині. Встановлено, що ЛПС бере участь у регуляції проникності
зовнішньої мембрани для різних речовин, захищаючи клітину від дії багатьох
детергентів, отрут, антибіотиків [3, 5, 8].
Грамнегативні бактерії мають
особливу будову клітинної оболонки, яка складається з зовнішньої мембрани,
внутрішньої цитоплазматичної мембрани та тонкого шару пептидоглікану.Двошарова
зовнішня мембрана має асиметричну будову. У зовнішньому шарі присутні в
основному білки та ліпополісахариди, до складу ж внутрішнього шару входять
білки та фосфоліпіди. Завдяки цьому наявність ліпополісахаридів є характерною
ознакою грамнегативних бактерій. Вони мають своєрідну будову молекули, яка
містить у своєму складі як гідрофільну, так і гідрофобнучастини, що надає
молекулі амфіфільних властивостей. Гідрофобною частиною молекула занурюється в
ліпідний шар зовнішньої мембрани, а гідрофільна частина знаходиться над
поверхнею мікробної клітини. Молекули ЛПС утворюють гідрофобний бар'єр, який, з
одного боку, обмежує надходження шкідливих речовин, таких як літичні ферменти,
отрути, деякі антибіотики, а з іншого – дають можливість бактеріям обійти
багато природних захисних факторіворганізму хазяїна[8].
Ліпополісахариди та білки-порини
зовнішньої мембрани зумовлюють її негативний заряд, що є характерним для
клітинної поверхні одноклітинних мікроорганізмів. Збільшення її гідрофільності
відбувається за рахунок вуглеводного шару, який утворений О-бічними ланцюгами
ліпополісахаридів. Клітинна поверхня гідрофільна навіть у тому випадку, якщо їх
ліпополісахарид не містить О-бічних ланцюгів, але подальше збільшення
гідрофільності корелює з послідовним підвищенням вмісту олігосахаридних одиниць
в О-специфічних ланцюгах.Це, очевидно, дає можливість клітинам уникнути
фагоцитозу, що необхідно для виживання патогенних бактерій в організмі
господаря, а також для вільноживучих мікроорганізмів.
Так, структура О-специфічних
бічних ланцюгів ліпополісахаридів різноманітна, тому виникнення нової
поверхневої структури, до якої імунна система тваринного організму не
пристосувалася, дає значну перевагу патогенним бактеріям.Поверхневі білки
зовнішньої мембрани, так звані порини, також дуже непостійні і, можливо,
володіють видовою специфічністю[3, 5, 8].
ЛПС (рис. 1), отриманий з
представників різних груп грамнегативних бактерій, як правило, складається з
гідрофільного гетерополісахариду, в якому розрізняють О-специфічний ланцюг і
коровий олігосахарид, та ковалентно приєднаного до нього ліпідного компонента –
ліпіду А. Така структура ЛПС – S-форма – продукується гладкими бактеріальними
фенотипами, представниками Enterobacteriaceae
і Pseudomonodaceae. Мікроорганізми,
які є штучно отриманими або природними R-мутантами (шорсткуваті фенотипи
багатьох видів Acinetobacter, Bordetella, Bacteroides, Neisseriae),
синтезують R-ЛПС, в молекулі яких відсутній О-полісахаридний фрагмент.
Більшість штамів «дикого типу»бактерій продукує S-, R-, а також перехідні
SR-форми ЛПС. У залежності від того, в S-або R-формі ЛПС присутній в клітинах,
застосовують різні методи їх виділення і очищення[8].
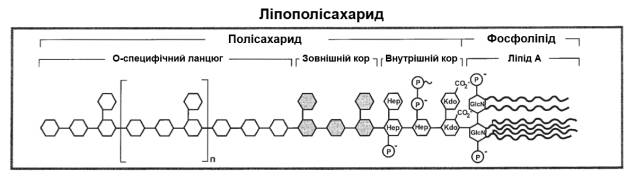
Рис. 1 Схема будови
ліпополісахариду
Ліпід А – компонент ліпополісахариду
У більшості вивчених
бактерій ліпід А являє собою фосфорильований дисахарид, побудований з двох
залишків глюкозаміну, які несуть залишки вищих жирних кислот, що приєднуються
ефірним чи амідним зв’язком. Ці залишки, що знаходяться по один бік
дисахаридної основи, орієнтовані перпендикулярно зовнішній мембрані та
утворюють компактний зовнішній шар, який утримується за рахунок гідрофобних
взаємодій з внутрішнім фосфоліпідним шаром мембрани[9]. Загальна риса ліпідів А
у тому, що вони містять дві вільні гідроксильні групи в положеннях 4 і 6'.
Остання гідроксильна група звільняється в ліпіді А тільки після кислотної
деградації ЛПС, що приводить до відщеплення полісахариду. Ця гідроксильна група
є місцем приєднання полісахарідного ланцюга через кетозидний зв'язок КДО до
ліпіду А [1].
Ліпід А – це найбільш
консервативна частина макромолекули ЛПС. Різноманітність будови ліпіду А в
різних бактерій визначається входженням до його складу різних жирних кислот.
Він визначає ендотоксичну активність макромолекули ЛПС і відповідає майже за
всі типи біологічної активності її та мікробної клітини в цілому. Він несе
відповідальність за цілий ряд патофізіологічних процесів, що викликаються ЛПС в
організмі ссавців, зокрема за його токсичні властивості, такі, як летальна
токсичність та пірогенність. Крім того, ліпід А має ад'ювантну і мітогенну
активність, стимулює проліферацію і секрецію імуноглобулінів, індукує утворення
лімфокінів, посилює фагоцитоз, активуючи комплемент і макрофаги[5]. Однозначні
докази ендотоксичних властивостей ліпіду А в макромолекулі було отримано після
порівняльного вивчення нативного ліпіду А E.coli, його синтетичного
аналога – сполуки 506 та її фрагментів – сполук 406 і Е5531.У результаті цих
досліджень було визначено мінімальні вимоги до структури біологічно активної
молекули ліпіду А. Для того, щоб стимулювати продукцію цитокінів вона має
містити два залишкигексозамінів, дві фосфатні групи і шість жирних кислот. Це
біологічно найбільш активна форма молекули ліпіду А. Ліпіди А, які не містять
хоча б одного з указаних фрагментів, є менш активними або й зовсім не активними
щодо стимуляції продукції цитокінів[8,11].
Кор – функціонально важлива ділянка ліпополісахарида
Кор представляє собою
кислий олігосахарид, пов'язаний безпосередньо з ліпідом А. Він присутній в ЛПС
всіх вивчених грамнегативних бактерій незалежно від того, чи є в молекулі
приєднаний до кору О-антигенний полісахаридний ланцюг (S- форма ЛПС) або
вуглеводна частина ЛПС обмежується тільки кором (R-форма). Єдиним структурним елементом,
який присутній у всіх ЛПС, незалежно від їх бактеріального походження, є
2-кето-3-дезоксіоктонова кислота (КДО) (від одного до трьох залишків) або її
похідні. Бактерії з дефектом у біосинтезі КДО не є життєздатними. Це вказує на
те, що КДО (як і ЛПС в цілому) є необхідною для структурної та функціональної
цілісності бактеріальної клітини. Незважаючи на те, що в ЛПС ряду бактерій
присутні 3 молекули КДО, для виживання грамнегативних бактерій достатньо тільки
одного залишку КДО [2, 8].
Коровий олігосахарид
умовно поділяють на дві структурні ділянки: внутрішній кор, або КДО-гептозна
ділянка, безпосередньо зв'язана з ліпідом А кетозидним зв'язком, та зовнішній
кор, або гексозна ділянка, ковалентно зв'язана з О-ланцюгом[8, 9].
КДО-гептозна ділянка –
специфічна субструктура в макромолекулі ЛПС, яка включає КДО і гептозу і
вважається найбільш консервативною її частиною. Роль КДО – забезпечити
кислотолабільний зв'язок між ліпідом А і полісахаридним ланцюгом у
глікополімері [6].Гексозна, або зовнішня, ділянка кору значно відрізняється від
внутрішньої. Набір цукрів, які в ній зустрічаються, набагато ширший, ніж у
внутрішньому корі. Основними її компонентами є такі широко розповсюджені цукри,
як глюкоза, галактоза, глюкозамін, галактозамін. Але останнім часом у складі
цієї ділянки макромолекули ЛПС ідентифіковано також багато унікальних цукрі
[8].
Ліпід А-КДО – мінімальна
структура, яка може забезпечити потреби бактерії, необхідні для організації й
функціонування зовнішньої мембрани. Завдяки наявності в макромолекулі ЛПС
КДО-гептозної ділянки, в якій розміщені полярні замісники, кор є основним
сайтом зв'язування катіонів. Взаємодія ж ЛПС з бівалентними катіонами необхідна
для збірки і підтримання нормальної структурної організації зовнішньої мембрани
грамнегативних бактерій, яка б забезпечувала їхній ріст та виживання [2].
Як і S-ЛПС, R-ЛПС виявляє
імуногенністьі при введенні в організм теплокровних стимулює продукцію
специфічних антитіл. Консервативність цієї частини макромолекули ЛПС зумовлює
наявність у корі епітопів, спільних для таксонів високих рангів. Наприклад,
отримано моноклональне антитіло, специфічне до роду Klebsiella. Причому антигенно активними є периферійні ділянки кору.
Так, встановлено, що всі доступні для запуску продукування і специфічного
зв'язування епітопи в корі ЛПС Salmonella
пов'язані із залишками, що створюють розгалуження основного полісахаридного
ланцюга. Багато штамів втратили здатність до синтезу S-ЛПС й існуть виключно в
R-формі, в тому числі і у природних умовах. Серед них такіпатогени, як Neisseriameningitis, N. gonorrhoeae, Н.
influenzae, Bordetellapertussis[4, 8].
ЗаголовокпроО-цепи
О-специфічні полісахариди
зазвичай характеризуються регулярною структурою, побудованою з повторюваних
олігосахаридних одиниць, що визначається способом їх біосинтезу, в якому раніше
утворені олігосахаридні блоки переносяться на зростаючий полісахаридний ланцюг.
У деяких випадках біосинтетичний шлях відрізняється від звичайного і тоді
моносахариди приєднуються до ланцюга послідовно один за іншим. Компоненти
О-ланцюга включають нейтральні моносахариди у піранозній і фуранозній формі
(гексози, пентози, дезокси-і О-метильні похідні) і заряджені моносахариди
(аміногексози і амінопентози, гексуронові і гексозаміноуронові кислоти), які
несуть ряд засмісників, таких як аміноацільні, фосфорильні, гліцерільні,
лактільні і ацетильні групи. Найбільш великий моносахарид містить 10 атомів
вуглецю. В останні роки в складі О-полісахаридів виявлені амінокислоти: серин,
треонін і аланін [7].
В основному
О-полісахариди представлені гетерополісахаридами, що містять до восьми різних
залишків моносахаридів. Природа, послідовність, аномерна конфігурація, тип
зв'язку і тип заміщення індивідуальних моносахаридних залишків всередині
повторюваної одиниці є характерними і унікальними для даного ЛПС і вихідного
штаму [8].
О-специфічний ланцюг
визначає серологічну специфічність ЛПС. Імуногенні і О-антигенні властивості
ЛПС визначаються так званими про-факторами, хімічна структура яких у багатьох
випадках встановлена. У ряді випадків встановлена кореляція між структурою і
серологічними властивостями ЛПС. З метою обдурити імунну систему хазяїна
бактерія в процесі еволюції змінює склад і структуру О-ланцюгів, що призводить
до розвитку нових О-специфічних
властивостей клітинної поверхні. Необхідний для росту і розмноження бактерій
ліпід А з приєднаною до нього зоною внутрішнього кора виявляються недоступними
для впізнавання клітинами господаря. Таким чином, О-специфічні ланцюги можуть
захищати бактерії від фагоцитозу і бактерицидної дії сироватки.
Заключення
Біосинтез
ліпополісахаридів грамнегативних бактерій протікає на цитоплазматичній мембрані
з наступним транспортом макромолекул до місця локалізації на зовнішній
мембрані.
Виступаючи основними
термостабільними антигенами і ендотоксинами мікробної клітини, ЛПС визначають
її серологічну специфічність, О-антигенність і значною мірою – характер
взаємодії з іншими мікроорганізмами та/або макроорганізмом у біоценозах, беруть
участь в інфекційному процесі в різних біологічних системах. Вивчення складу та
будови ЛПС сприяє розумінню на молекулярному рівні найскладніших біологічних
процесів, які відбуваються за їх участю[8, 10].
Література:
1.
Варбанец Л. Д. Структура, функция, биологическая
активностьэ ндотоксинов грамотрицательных бактерий / Л. Д. Варбанец, Н.В.
Винарская // Совр. проблемы токсикологии. – 2002. – №1. – C. 33-45.
2.
Варбанец Л. Д. Эндотоксины грамотрицательных
бактерий: структура и биологическая роль / Л. Д. Варбанец // Мікробіологічний
журнал. – 1994. – Т. 56, № 3. – С.227-235.
3.
Вінарська Н.В. Залежність біологічної активності
ліпополісахаридів Ralstonia solanacearum
(Smith, 1896) від їх складу та структури: автореф. дис. на здобуття наук.
ступеня. канд. біол. наук: спец. 03.00.07 «Мікробіологія» / Н. В. Вінарська –
К., 2002. – 20 с.
4.
Здоровенко Э.Л. Биологическая активность
липополисахаридов грамотрицательных бактерий/ Э. Л. Здоровенко, В. К. Позур, М.
Е. Кучеренко // Биополимеры и клетка. – 2000. – Т. 16, № 1. –С. 5-15.
5.
Книрель Ю. А. Строение липополисахаридов
грамотрицательных бактерий. I. Общая характеристика липополисахаридов и
строение липида А / Ю. А. Книрель, Н. К. Кочетков // Биохимия. – 1993. – Т. 58,
№ 2. – С. 166-181.
6.
Книрель Ю. А. Строение липополисахаридов
грамотрицательных бактерий. II. Структура кора / Ю. А. Книрель, Н. К. Кочетков
// Биохимия. – 1993. – Т. 58, № 2. – С. 182-201.
7.
Книрель Ю. А.
Строение липополисахаридов грамотрицательных бактерий. III. Структура
О-специфических полисахаридов / Ю. А. Книрель, Н.К. Кочетков // Биохимия.
– 1993. – Т. 58, № 2. – С. 202-217.
8.
Структура і біологічна активність бактеріальних
біополімерів / В. К. Позур, Д. В. Колибо, Л. Д. Варбанець [та ін.]. – К.:
Видавничо-поліграфічний центр “Київський університет”, 2003. – 305 с.
9.
Bacterial toxins: methods and protocols / edited by
Otto Holst. – Totowa, New Jersey: Humana Press Inc., 2000. – Vol. 145 – 2000. –
392 p.
10. Raetz
C.R. Biochemistry of endotoxins / C.R. Raetz // Ann. rev. biochem. – 1990 – Vol. 59. – Р. 129-170.
11. Synthetic
and natural Escherichia coli free lipid
A express identical endotoxic activities / C. Galanos, O. Lüderitz, E. T.
Rietschel [etal.]// Eur. J. biochem. – 1985. – Vol.
148, № 1. – P. 1-5.