Медицина/8. Морфология
к.м.н. Купша
Е.И., доцент Бондаренко В.В.
Государственное учереждение “Крымский государственный медицинский университет
имени С.И. Георгиевского”, АР Крым, Украина
Митофагия в гепатоцитах мышей при поступлении в организм малых доз ацетата
свинца.
В клетках млекопитающих выделяют три вида аутофагии:
макроаутофагия, микроаутофагия и шаперон-медиаторная аутофагия, которые все
приводят к деградации цитозольного компонента в лизосомах [1]. Макроаутофагия
включает образование двойной мембранной структуры в пределах цитоплазмы которая
охватывает и изолирует органеллы или цитоплазму формируя аутофагосому. После
слияния с лизосомой внешняя мембрана аутофагосомы становится частью лизосомы, а
внутренняя мембрана и содержимое деградирует в пределах лизосомы [2]. Удаление
митохондрий, содержащих поврежденные компоненты, осуществляется через аутофагию
(митофагию). Митофагия призвана обеспечить баланс в биогенезе, обеспечивая
соответствие энергетической потребности тканей. Защитный эффект митофагии выражается
в элиминации поврежденных, агрегировавших совокупностей протеинов и патогенов,
избыточных митохондрий и, таким образом, снижении уровня оксидации и проапоптозных
молекул (подобных цитохрому С) [3, 4]. Митофагию подразделяют на два класса: а)
«поддерживающая» митофагия и б) митофагия ассоциированная со стрессом или
клеточной дифференцировкой. Первый класс включает активацию митофагии в ответ
на повреждение митохондрий, приводя к целенаправленному удалению и деградации
этих органелл. Второй класс затрагивает нормальные физиологические условия при
которых митофагия играет роль в процессах клеточной дифференцировки. Таким
образом, жизнеобеспечивающее знфчение митофагии включает ряд цитопротекторных результатов
– уменьшение АФК, производимых поврежденными митохондриями и исключение
необходимости поддержания неэффективных органелл [5].
Установлено,
то аутофагию индуцирует оксидативный стресс [6]. Известно, что введение
субтоксических доз свинца вызывает критическое возрастание АФК и оксидативный
стресс в печени крыс [7].
Свинцовая интоксикация моделировалась нами на 6 самцах
белых мышей линии balb/c, которым, начиная с 1-месячного возраста, ежедневно
перорально вводили ацетат свинца в дозе 10 мг/кг/сутки в течение 30 суток.
При электронномикроскопическом исследовании нами была визуализирована
митофагия в гепатоцитах с морфологическими признаками высокой функциональной
активности клеток паренхимы (рис. 1).
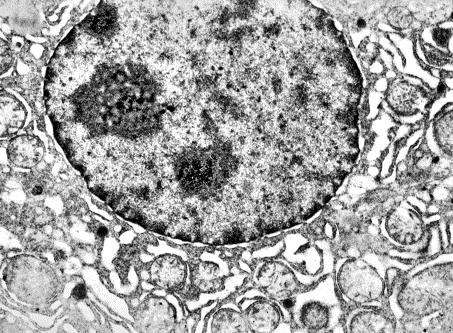

Рис.
1. Митофагия с участием ГрЭПС в околоядерной зоне – сайте митохондриального
генеза. Электронная микрофотография. Ув. х 10000.
Ядра таких гепатоцитов крупные, округлой формы, с неглубокими инвагинациями кариолеммы. В них преобладает транскрипционноактивный эухроматин. Гетерохроматин представлен в умеренных количествах и распределяется по всей кариоплазме. Небольшое его количество равномерно и типично локализовано под кариолеммой. Ядрышек 1-2, в некоторых клетках до 4-х. Ядрышки – продуценты рибосомальных субъединиц – локализуются эксцентрично, имеют округлую форму и обычную электронную плотность. В мелких ядрышках лучше выражен гранулярный, а в крупных – фибриллярный компоненты. Ядерные поры увеличены и четко контурированы, в местах их локализации примембранный гетерохроматин отсутствует. Большая часть поровых комплексов электронноплотная. К наружной ядерной мембране по всему периметру прилежит ГрЭПС. Промежутки между ее цистернами сильно расширены. Сами цистерны гипертрофированы, заполнены электронноплотным содержимым и интимно контактируют с митохондриями, в отдельных случаях сливаясь с их наружной мембраной. Такое расположение подтверждает данные литературы о превалирующей роли ГрЭПС в инициации формирования мембраны транспортной фагосомы [8, 9].
Учитывая распределение свинца в гепатоцитах и его
преимущественное расположение в митохондриях [10], логично заключить, что митофагия
играет роль в адаптации количества и качества митохондрий к новым условиям,
чтобы исключить поврежденные, дисфункциональные и лишние митохондрии в рамках
нормы реакции, что согласуется с мнением ряда исследователей [11].
Литература:
1. Danielle
Glick. Autophagy: cellular and molecular mechanisms / Danielle Glick, Sandra Barth, Kay F. Macleod // J
Pathol. – 2010. – Vol.221, №1 – P.3–12.
2. Harmeet Malhi. Hepatocyte Death: A Clear and
Present Danger / Harmeet Malhi, Maria Eugenia Guicciardi, and Gregory J. Gores
// Physiol Rev. – 2010. – Vol.90
– P.1165–1194.
3. Gottlieb RA. Autophagy in health and
disease: 5. Mitophagy as a way of life / Gottlieb RA, Carreira RS. // Am J
Physiol Cell Physiol – 2010. – Vol.299 – P.203–210.
4. Kevin Moreau. Cytoprotective roles for
autophagy / Kevin Moreau, Shouqing Luo and David C Rubinsztein // Current
Opinion in Cell Biology – 2010. – Vol. 22 – P. 206–211.
5. Scott
J. Goldman. Autophagy and the degradation of mitochondria / Scott J. Goldman,
Robert Taylor, Yong Zhang, and Shengkan Jin. // Mitochondrion – 2010. – Vol.10 №4 – P.309–315.
6. Chen Y. Superoxide is the major reactive
oxygen species regulating autophagy / Chen Y, Azad MB, Gibson SB. // Cell Death
Differ – 2009. – Vol.16 – 1040–1052.
7. Saxena G. Lead-induced oxidative stress and
hematological alterations and their response to combined administration of
calcium disodium EDTA with a thiol chelator in rats / G. Saxena, S.J. Flora //
J Biochem Mol Toxicol. – 2004. – Vol.18, №4. – Р.221-233.
8. Hayashi-Nishino M. A subdomain of the
endoplasmic reticulum forms a cradle for autophagosome formation / Hayashi-Nishino
M, Fujita N, Noda T, Yamaguchi A, Yoshimori T, Yamamoto A // Nat Cell Biol. – 2009.
– Vol.11 – P.1433–1437.
9. Yla-Anttila P. 3D tomography reveals
connections between the phagophore and endoplasmic reticulum / Yla-Anttila P,
Vihinen H, Jokitalo E, Eskelinen EL // Autophagy – 2009. – Vol.5 – P.1180–1185.
10.
Луговской С.П. Накопление и распределение свинца в ультраструктурах гепатоцитов
крыс / С.П. Луговской // Соврем. пробл. токсикологии – 2004. – № 1 – С.35-37.
11. Ingrid Bhatia-Kiśśová.
Mitophagy inyeast: actors and physiological roles /Ingrid Bhatia-Kiśśová,
Nadine Camougrand // FEMS Yeast Res. – 2010. – Vol.10 – P.1023–1034.