Битемирова У., Балабеков Б.Ч.
ЮКГУ имени М.О. Ауезова, г. Шымкент, Казахстан
Математическая модель образования структур
в сообществе микроорганизмов
Пространственно-временная
самоорганизация микробиологических систем часто возникает как в естественной
среде, так и в лабораторных опытах [1]. Для объяснения образования
пространственной неоднородности в одних случаях предлагалось учитывать внешние
химические градиенты, в других случаях необходимо принимать во внимание либо
хемотаксис клеток, либо генетический триггер. Но также описаны опыты, в которых
ни одна из перечисленных причин не дает адекватного объяснения. В этих опытах
микроорганизмы культивировались на питательном полужидком агаре в плоской
чашке. При таких условиях подвижность микроорганизмов такова, что колония,
инициированная одной клеткой, разрасталась почти по всей чашке за время
инкубации. Внешне колония представляла собой картину чередующихся
концентрических колец высокой и низкой плотности клеток. Пространственный
период и полный размер колонии можно было изменить, варьируя температуру
инкубации или плотность агара. В некоторых случаях кольца могли распадаться на
регулярно расположенные пятна приблизительно одинакового размера. Расположение
пятен визуально соответствует гексагональной симметрии. В данной системе нет
противоположно направленных химических градиентов. Хемотаксис также не играет
важной роли, так как для мутантных штаммов использованных микроорганизмов,
лишенных способности к хемотаксису, наблюдались кольцевые картины. Опытные
данные подтверждают, что образуемые кольцевые образования являются
неподвижными. Это подсказывает о том, что для разработки модели можно
использовать известную теорию диссипативных структур.
В данной работе предлагается
математическая модель образования пространственных структур в колонии
микроорганизмов. Наблюдаемый неоднородный рост колонии, по-видимому, обусловлен
взаимодействием клеток. Разумно предположить, что это взаимодействие
опосредовано диффузионно подвижным химическим фактором, например, каким-либо продуктом
их клеточного метаболизма. Теория диссипативных структур подсказывает, что для
создания пространственной неоднородности в рассматриваемой системе этот гипотетический
фактор должен действовать как ингибитор роста клеток. Таким образом, наша
модель будет описывать изменения концентраций микроорганизмов и ингибитора.
Далее будет предполагаться, что производство ингибитора прямо пропорционально
концентрации клеток, т.е. каждая клетка вырабатывает ингибитор с постоянной
скоростью. Также считаем, что ингибитор распадается с постоянной скоростью,
т.е. имеет конечное время существования. Клетки в колонии могут двигаться
хаотично, что может быть описано через эффективную диффузию, причем
диффузионная подвижность ингибитора считается более высокой, чем диффузионная
подвижность клеток. Для роста клеток делается следующее допущение о том, что он
подчиняется известному биологическому закону, гласящему, что существует
некоторая благоприятная плотность популяции для выживания и оптимального роста
сообщества. То есть, как малочисленность (малая плотность), так и
перенаселенность (большая плотность) могут оказывать лимитирующее влияние на
рост колонии. Общий характер функции роста популяции от плотности должен иметь
колоколообразный вид. Этот принцип был использован при создании модели,
описывающей систему типа хищник-жертва для планктона, где зоопланктон выступал
в роли хищника, а фитопланктон играл роль жертвы. Наконец принимается, что рост
клеток уменьшается пропорционально концентрации ингибитора. С учетом всех сделанных
предположений можно записать систему уравнений для изменения концентраций
микроорганизмов (В) и ингибитора (Н) в плоском случае в размерных величинах
следующим образом:
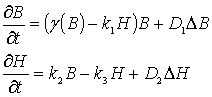 , (1)
, (1)
где D - двумерный лапласиан,
функция g(B) имеет
вид:
![]() .
.
D1, D2 – размерные коэффициенты диффузии клеток и
ингибитора соответственно, k1, k2, k3 –
размерные коэффициенты системы, ![]() - положительные
параметры, характеризующие функцию роста микроорганизмов,
- положительные
параметры, характеризующие функцию роста микроорганизмов, ![]() - точка равновесия
системы. Система (1) имеет однородное нетривиальное стационарное решение:
- точка равновесия
системы. Система (1) имеет однородное нетривиальное стационарное решение:
![]() ,
, ![]() . (2)
. (2)
Перейдем к безразмерным
величинам, вводя обозначения:
![]() (3)
(3)
Учитывая (2) и (3) получим
систему уравнений:
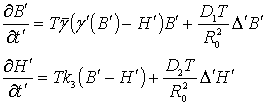 (4)
(4)
если примем, что ![]()
и обозначим 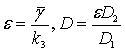 ,
,
опуская штрихи получим
безразмерную систему в виде:
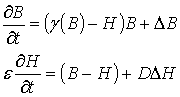 (5)
(5)
Здесь функция g(B) имеет
вид:
![]() (6)
(6)
Таким образом, получили
безразмерную модель (5), (6) для образования пространственных структур в
системе клетки – ингибитор, которая содержит следующие параметры: a, b, e, D. Если принять быструю
кинетику ингибитора, то величина параметра e будет малой. Это позволит
эффективно исключить второе уравнение системы и в целом облегчит решение
задачи.
Литература
1. Wimpenny J.W.T., Lovitt R.W., Coombs J.P. Microbes in their natural
enviroments. – Cambridge Univ. Press, 1983.