Щеколова А.С., к.б.н. Квач С.В., д.б.н.
Зинченко А.И.*
Институт
микробиологии НАН Беларуси, Минск
*Международный
государственный экологический университет
имени А.Д.
Сахарова, Минск
МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКАЯ ХАРАКТЕРИСТИКА ПЛАЗМИДЫ, ОБОГАЩЕННОЙ CpG-МОТИВАМИ
CpG-мотив представляет собой
CpG-динуклеотид вместе с одним или двумя нуклеотидами на его 3'- и 5'-концах.
Неметилированные CpG-мотивы преобладают в ДНК прокариот. Эти участки ДНК
распознаются иммунной системой позвоночных как «сигнал тревоги», активируя как
врожденную, так и адаптивную ветви иммунитета и во много раз усиливая ответ
организма на антигены, т.е. проявляют адъювантную активность [1].
Одним из способов получения таких адьювантов является создание
плазмид, обогащенных CpG-мотивами с
их последующим клонированием в бактериальных клетках [2]. В научной литературе
имеются данные о том, что с увеличением числа копий CpG-мотивов, встроенных в плазмиду, повышается и
биологическая активность такой CpG-ДНК [3].
Таким образом, при создании CpG-плазмид
большое значение имеет определение точного числа встроенных CpG-мотивов.
Ранее в лаборатории молекулярной
биотехнологии Института микробиологии НАН Беларуси была сконструирована
плазмида pCpG-KH11, обогащенная множественными повторами GTCGTT-CpG-мотива,
который согласно литературным данным обладает стимулирующей активностью по
отношению к иммунной системе человека [4].
Исходя из описанного выше, целью настоящей
работы явилась молекулярно-генетическая характеристика плазмиды pCpG-KH11 для
определения количества встроенных в нее CpG-мотивов.
Объекты
и методы исследования. В работе
использована плазмида pCpG-KH11, сконструированная на основе вектора pXcmkn-12 (Cloning
vector collection,
Япония) и содержащая множественные повторы GTCGTT-мотива.
ПЦР-анализ плазмиды pCpG-KH11 на наличие
встроенных CpG-мотивов проводили в 50 мкл
реакционной смеси, содержащей 10 нг целевой плазмиды, 67 мМ Трис-HCl (pH 8,3), 17 мМ (NH4)2SO4, 2 мМ MgCl2, 0,02%
Твин-20, смесь четырех канонических дНТФ (каждого в концентрации 0,2 мМ),
по 5 пмоль праймеров, ограничивающих клонированный фрагмент, и 1 ед Taq-ДНК-полимеразы.
ПЦР проводили по следующей программе: этап начальной денатурации 2 мин 95°С; 25
циклов амплификации: 30 с при 95°С; 15 с при 55°С; 1 мин при 72°С; финальная
элонгация 2 мин при 72°С.
Рестрикционный анализ плазмиды проводили
путем обработки вектора CpG-KH11 смесью рестриктаз NdeI и EcoRI с
последующим инкубированием продуктов реакции с рестриктазой HindIII.
Полученные в результате ПЦР-амплификации и
рестрикции pCpG-KH11 фрагменты разделяли с использованием электрофореза
в 1% агарозном геле.
Результаты и обсуждение. С целью определения точного числа CpG-мотивов, клонированных в плазмиде pCpG-KH11 проведен
ПЦР- и рестрикционный анализ целевого вектора.
На первом этапе методом ПЦР амплифицирован
фрагмент вектора, содержащий клонированные CpG-мотивы и участки вектора, ограничивающие
клонированную «вставку». ПЦР-амплификат разделяли в 1% агарозном геле (рис. 1,
дорожка 1). Размер полученного фрагмента, определенный с иcпользованием пакета программ TotalLab TL120,
составил 1100 п.о. Без учета длины фланкирующих участков (121 п.о.), размер
клонированного фрагмента составил 979 п.о.
Рестрикционный анализ
проводили с помощью рестриктаз NdeI и EcoRI, сайты узнавания которых
ограничивают клонированную «вставку». При этом полученный фрагмент содержал
также участки вектора, фланкирующие клонированную «вставку». Длина вырезанного
фрагмента, определенная с иcпользованием
пакета программ TotalLab TL120, составила 1250 п.о. (рис. 1, дорожка 4).
Учитывая, что суммарная длина фланкирующих участков составляет 286 п.о., то
согласно рестрикционному анализу клонированная «вставка» имеет длину 964 п.о.
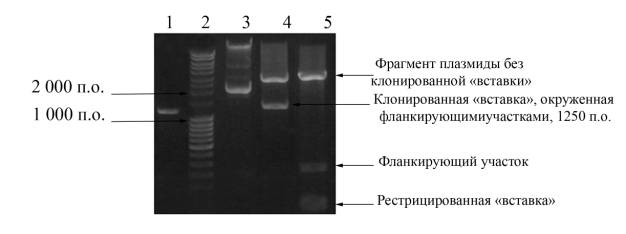 Рис. 1 –
Электрофореграмма продуктов ПЦР-амплификации и рестрикции плазмиды рCpG-KH11
Рис. 1 –
Электрофореграмма продуктов ПЦР-амплификации и рестрикции плазмиды рCpG-KH11
1 – продукт ПЦР-амплификации плазмиды
рCpG-KH11; 2 –маркеры молекулярных масс фрагментов ДНК; 3 – очищенная плазмида
рCpG-KH11; 4 – фрагменты ДНК, полученные после обработки рCpG-KH11 смесью
рестриктаз NdeI и EcoRI; 5 – фрагменты ДНК, полученные после обработки
рCpG-KH11 смесью рестриктаз NdeI, EcoRI и HindIII
Клонированная «вставка» состоит из
тандемных повторов олигонуклеотидов, содержащих по 4 GTCGTT-мотива и разделенных сайтами узнавания рестриктазы HindIII. Содержание CpG-мотивов в клонированной вставке рассчитывали
следующим образом:
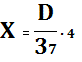 , где:
, где:
D – размер клонированного фрагмента, определенного с
помощью рестрикционного или ПЦР-анализа (п.о.);
37 – размер одного тандемного повтора CpG-мотивов (п.о.);
4 – количество CpG-мотивов в одном тандемном повторе.
Таким образом, содержание CpG-мотивов в плазмиде рCpG-KH11 составило 104 повтора,
что превосходит аналогичный показатель известных в настоящее время
плазмид-аналогов [5].
Наличие CpG-мотивов в клонированной вставке подтверждали путем
дополнительной обработки продуктов рестрикционного анализа эндонуклеазой HindIII, сайты узнавания которой
находятся внутри клонированного фрагмента (рис. 1, дорожка 5). По деградации
клонированной «вставки» можно сделать вывод о том, что она действительно
содержит тандемные повторы CpG-мотивов.
В настоящей работе с помощью
ПЦР- и рестрикционного анализа проведена молекулярно-генетическая
характеристика вектора рCpG-KH11.
Показано, что данный вектор содержит 104 повтора иммуностимулирующего GTCGTT CpG-мотива,
что на сегодняшний день является наибольшим по сравнению с известными CpG-плазмидами.
Литература:
1.
Krieg, A.M. CpG still rocks! Update on an accidental drug / A.M. Krieg
// Nucleic Acid Ther. – 2012. – Vol. 22, N 2. – P. 77–89.
2.
Adjuvant effect of multi-CpG motifs on an HIV-1 DNA vaccine /
Y. Kojima [et al.] // Vaccine. – 2002. – Vol. 20, N 23–24. – P. 2857–2865.
3.
The introduction of multi-copy CpG motifs into an antiviral DNA vaccine
strongly up-regulates its immunogenicity in fish / S. Martinez-Alonso
[et al.] // Vaccine. – 2011. – Vol. 29. – P. 1289–1296.
4.
Krieg, A.M. CpG motifs in bacterial DNA and their immune effects /
A.M. Krieg // Annu. Rev. Immunol. – 2002. – Vol. 20. – P. 709–760.
5.
Chen, Y. Construction of a recombinant plasmid containing multi-copy CpG
motifs and its effects on the innate immune responses of aquatic animals /
Y. Chen, L.X. Xiang, J.Z. Shao // Fish Shellfish Immunol. – 2007. – Vol. 23, N 3. – P.
589–600.