*112960*
НОРМЫ
РАСХОДА ЭНЕРГИИ ПРИ ДВИЖЕНИИ ПО ЗЕМЛЕ (4-й закон
механики)
УДК
531.4 В.И. Голубев
Основной принцип в природе – это
экономия энергии. В чем это
выражается? Все живые существа на
планете передвигаются с довольно строгим соблюдением закона энергии Е=m·v2/2.
Масса тела должна удовлетворять
требованию m =2E/v2 , т.е. экономия энергии
эквивалентна уменьшению массы.
Знать это недостаточно, это
нужно понимать, с этим надо жить, чувствовать это во всех ситуациях. Биологи обошли энергетическую формулу
стороной, никто не уделил ей серьезного внимания. Из-за большого
диапазона
массы тел и их
размеров, они сразу перешли к формуле
Р=aM b [1,2].
Эта аллометрическая формула позволяет большой диапазон значений сузить и
поместить эти значения на одном листе с
логарифмической шкалой. Она была введена Клайбером в 1932г, т.е. во время увлечения логарифмической линейкой, и позволяла укладывать на одном графике
самые разные параметры биологических тел и получать линейную зависимость в
логарифмической системе координат. Биологи увлеклись этой формулой и принялись
исследовать все свойства живой материи,
а также математическое описание движения биологических тел и их физиологические
особенности. В итоге многочисленных
исследований было установлено, что на краях диапазона размеров тел живого мира
величина основного метаболизма отличалась от среднего уровня, и это мешало
сформулировать основной закон природы,
а именно закон экономии энергии всех биологических видов. Оставалось неясным,
что привело к минимальному расходу энергии на основной метаболизм и на
движение. Всестороннее изучение основного метаболизма отвлекло внимание ученых
от измерения энергии на движение в номинальных условиях.
Внутренние процессы в организме
определены как основной обмен, это неотъемлемая часть существования белковых тел.
Но борьба за выживание требует крепкого
тела, быстрого движения его в предназначенном для него пространстве, это ведет к увеличению отдельных частей тела
и в итоге всей массы. Каждое
существо было разработано для определенного пространства, в котором оно
двигается и добывает себе энергию для дальнейшего движения, это дает ему одну
степень свободы в рамках этого пространства (поле, лес, огород, река, море,
горы и т.д.) и определяет его габариты
и вес. Здесь ему надо двигаться со скоростью, которая определена борьбой внутри
этого пространства, и иметь запас биологической энергии, которую оно добывает
для своего движения в данном ему пространстве. Запас энергии необходим также
для борьбы с другими видами, которые внедряются в это пространство.
Определение величины этой номинальной скорости можно
осуществить несколькими способами. Первый состоит в том, что мы исходим из
основного принципа природы – экономии энергии.
Инверсия в сознании
энергетической формулы, которая сформулирована на основании 2-го закона
Ньютона, дает следующие
зависимости V (m) = ![]() при фиксированной E, это гиперболическая зависимость, и V(E) =
при фиксированной E, это гиперболическая зависимость, и V(E) = ![]() – это обратная квадратичная зависимость.
– это обратная квадратичная зависимость.
Эти зависимости целесообразно рассмотреть на совмещенном графике m – V –
E, рис.2б, где ось
ординат является общей, а ось абсцисс разделена на две части: m
и E. При таком подходе нам проще совместить математику вопроса с его философским
анализом, который превалирует над физикой взаимодействия массы с энергией. Это
объясняется тем, что между ними проходит сложная череда взаимодействия
биологических тел и их физиологических
особенностей: разное потребление
кислорода, рассеивание тепла через поверхность тела, размеры тела, важные при
взаимодействии его с другими телами в борьбе видов, и прочие биологические
параметры. А также взаимодействие
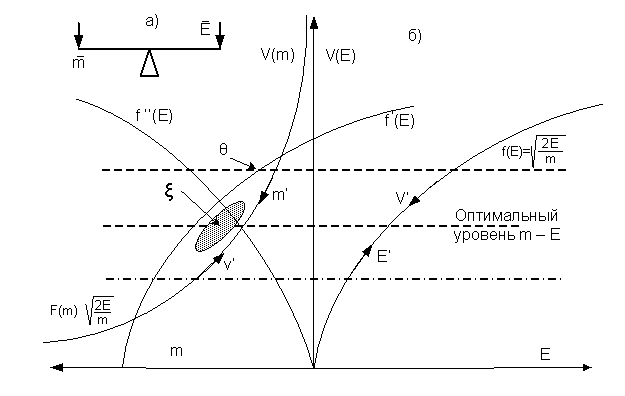 Рис.2
Графики поиска оптимального соотношения массы и энергии на передвижение.
Рис.2
Графики поиска оптимального соотношения массы и энергии на передвижение.
тел разных видов пространства: воды, воздуха, суши [2]. При таком
совмещении двух графиков мы получаем возможность найти минимум энергии
для данной массы при выборе оптимальной скорости.
Для каждого
биологического вида m и Е
находятся на весах выбора средних величин m' и Е', рис.2а, и весы эти
приходят в положение равновесия, когда масса переходит в энергию, а энергия
определяет величину массы в пространстве существования данного вида. Вот где определяются размеры
тел животных, которые Шмидт-Нильсен [2] не смог обосновать, т.к. ответ находится не в поле
биологии, а в области физики и
философии. Мало оперировать только
аллометрической формулой, надо рассматривать биологический мир с позиций разных
наук. И учесть также, что главный
конструктор биологических тел хотел, чтобы они удовлетворяли потребностям
человека, а по размерам подходили к его
росту, чтобы они были удобными для
человека в пользовательском смысле, будь то лошадь, корова, собака, курица,
рыба, голубь и т.д. Если животные
(динозавры) не стыкуются по своим свойствам и размерам с человеком, и он не
может с ними жить и справиться простыми средствами, то их пришлось
ликвидировать.
Колебания величины энергии ведут к смещению скорости в точку оптимального расхода энергии для заданной массы, рис.1 и рис.2б. На
рис.2б стрелками показано смещение номинальной скорости и расхода энергии в
точку их оптимального соответствия. Эту точку легче найти, если
плоскость графика V(E) повернуть вокруг оси V и мы получим пересечение графика f' '(E) с графиком F(m). Это и есть оптимальная
величина массы для данного биологического вида. Но поиск ее идет
эволюционно, в зависимости от
величины g на нашей планете, количества кислорода в воздухе, pH воды и других факторов
природной среды.
Колебания скорости вдоль левого графика (показано
стрелками m' — v') вызывает соответствующее изменение
энергии для этого движения на правом графике
V' — E'. Момент количества движения на левом
графике должен быть равен расходу энергии ΔЕ
на правом графике. Эти колебания можно рассмотреть на весах массы и энергии,
рис.2а. Для каждого биологического тела
масса и энергия находятся на весах средних величин m' и E' для конкретного биологического
вида. И весы эволюции приводят их в
положение равновесия, когда масса в процессе работы переходит в энергию и эта
энергия определяет величину массы в
пространстве существования данного
вида. Заметное превращение массы в энергию дает перелет
птиц на большие расстояния, 1000 км и более, а также перелеты самолетов, когда
масса переходит в энергию, она при этом уменьшается на 30 – 40% [11,12].
То же самое получим, если сдвинуть кривую f(E) влево (2-й способ) до пересечения графиков. Так мы тоже получаем
соответствие между массой биологического тела и энергией для него. Но это уже
не точка, а зона соответствия между массой и энергией – ξ. Смещение точки оптимального соответствия
вдоль графика правой кривой V(E) = ![]() эквивалентно смещению точки равенства вдоль левой кривой V(m) =
эквивалентно смещению точки равенства вдоль левой кривой V(m) = ![]() . Константы
находятся из общего уравнения энергии:
E = m·V2/2 = соnst2·Vn2 , откуда сonst2 = m/2 и
сonst1
= m· Vn2 = 2E n.
. Константы
находятся из общего уравнения энергии:
E = m·V2/2 = соnst2·Vn2 , откуда сonst2 = m/2 и
сonst1
= m· Vn2 = 2E n.
Во втором случае кривые дают величину
энергии для данной массы при номинальной скорости. Величину этой скорости мы
определяем изначально путем обработки статистических наблюдений за поведением
конкретного животного. По этой скорости
получаем среднюю величину массы m' = сonst1/
v2n =
2E n / v2n и далее среднюю величину
энергии для нее E' = сonst2 · Vn2 . Такой подход со
стороны массы и энергии к определению размеров биологических тел и его
логический анализ дает ответ на поставленный в
[2] вопрос о размерах тела.
Можно ли считать это достаточным обоснованием, покажет время и следующая
статья, в которой рассматриваются не только пешие, но и плавающие и летающие
живые существа. В
зависимости от конструкции биологического тела, которая определяется условиями
его жизни и движения в рамках той степени свободы, которая ему
предопределена изначально, мы имеем
такие варианты превращений массы в энергию и наоборот.
1.Если масса велика, то среднестатистическая скорость мала, у тела
мало энергии.
2.Жизнь заставляет уменьшать массу, скорость растет, тело достигает успехов в естественном отборе, что требует увеличения запасов энергии. Масса переходит в энергию мышц.3.Если энергии много (точка θ на f '(E), рис. 2б), скорость растет, масса уменьшается. Если тело набирает массу – скорость падает. И эти колебания поиска оптимальных величин определены как естественный отбор.
Средний расход
энергии при быстром передвижении Еmax= m · V2max/2.
Тогда он возрастает в квадратичной зависимости от скорости и достигает
максимума Eимп на
предельной скорости для данного вида. Так природа естественного отбора
осуществляет поиск оптимального соответствия основных параметров биологического
тела в пространстве физических величин. Но нам важно знать изменение энергии в
единицу времени, т.е. мощность на движение массы, а это есть производная от
энергии по времени: dE/dt = d (m · Vn2 /2) /dt . _
Номинальная скорость достигается
достаточно малым расходом энергии, когда дыхание не требует дополнительного
потребления кислорода, потребление кислорода в этом случае совпадает с
потреблением его в состоянии покоя.
Возрастание потребления кислорода говорит об интенсификации двигательной
активности и увеличения энергетического расхода выше уровня основного обмена
[3]. Для лучшего понимания взаимной
связи массы, скорости и энергии можно рассмотреть пространственную модель связей между ними. Это третий способ рассмотрения. В плоскости
(Е, m)
мы имеем линейную зависимость, – энергия и масса тела линейно связаны между
собой. В плоскости (m,v) получаем гиперболу. В плоскости (Е, v) имеет место квадратичная
зависимость Е от v. В результате получилась трехмерная
модель, рис.3, которая в зависимости от параметров
представляет геометрический образ в виде треугольного параллелепипеда,
образованного плоскостями µ-β-σ и
µ'-β'-σ', который необходимо
представить, и это можно сделать. Если
прокрутить его в уме много раз во все стороны, то можно почувствовать, что все параметры надо уменьшать, из
соображений экономии энергии для заданной массы. Но с другой стороны, борьба
естественного отбора требует от каждого биологического тела максимальной
скорости, как в беге, так и в движениях. Эта борьба и экономия энергии ведет к
компромиссу между этими параметрами
[6,8,2, 21]. Борьба видов
требует максимума v для конкретного вида в координате его жизни и движения. Если
принять, что каждый вид имеет свою степень свободы, и движение данного вида
осуществляется в рамках этой степени свободы, то рассмотренные плоскости должны
сближаться. Здесь под степенью свободы понимается пространство, в рамках
которого данный биологический вид
добывает себе пищу-энергию.
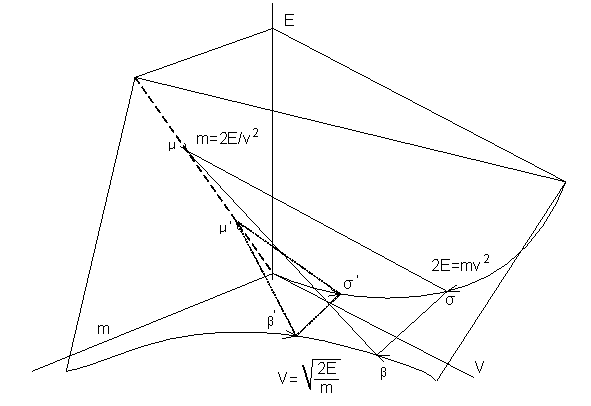
Рис.3 Пространственная
модель оптимального соответствия массы, энергии и скорости.
Чтобы иметь максимум v
надо иметь запас Е. Не в данный момент, а всегда, во всех статистически
наблюдаемых моментах времени.
Максимум v нужен в борьбе видов за выживание, чем быстрее двигается m, тем
больше шансов (вероятность) ему выжить в борьбе с другими m
с их собственными v. Причем не только в скорости
передвижения, но и в скорости действий.
Масса жестко связана с энергией, для скорости, фиксированной в данной координате. Т.к. энергию надо экономить,
то масса должна быть минимальной для фиксированных скоростей этого вида биологических тел, которую определяют все другие представители этого вида. Если мы перевернем главную формулу энергии,
то получим m = 2Е /v2
, откуда видно,
что для экономии энергии необходимо
иметь минимальную массу.
Компромисс в выборе m,
v
и Е заключен между
плоскостями µ-β-σ и µ'-β'-σ'.
Первая плоскость требует понижения этих величин, из соображений экономии энергии, вторая – увеличения
их, из соображений выживания в условиях борьбы видов. Оптимизация выбора приводит к тому, что эти плоскости сближаются и
в итоге сливаются в одну точку или
между ними остается малый разрыв, это зона ξ на рис.2б.
В свою очередь, скорость зависит от запаса энергии и от величины массы.
Рассмотрим еще один вариант расчета мощности движения по поверхности земли. Это движение по земле осуществляется с помощью ног,
которые шагают по земле. И двуногие, и четвероногие. В отличии от расчета Белецкого
и Неймарка [13,14] здесь
рассматривается плоская картина двуногой ходьбы без бокового наклона и движения
в гору. Мышцы ног передвигают тело так,
что оно при каждом шаге поднимается на высоту дуги Δh1,
рис.4. Движение происходит под
действием проекции силы мышц на горизонтальную плоскость, которая
уравновешивается силой трения ноги Fтр о поверхность почвы. Чтобы уменьшить
колебания тела, у ноги сделан коленный сгиб и изгиб ступни, в результате чего в
среднем положении и в начале шага нога укорачивается, подъем тела уменьшается
до величины Δh2
и энергия на передвижение тоже уменьшается, одновременно достигается
амортизация подъема и опускания тела, уменьшается вибрация внутренних
органов тела. 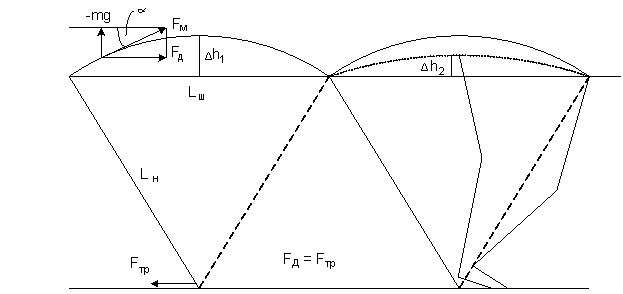
Рис.4.
К расчету энергии
передвижения шагающего тела
При передвижении за счет ног тело совершает
колебательное движение вверх-вниз, при подъеме тела энергия тратится на
преодоление силы тяжести. Сила мышц FМ
ноги
проектируется на горизонтальную плоскость, где она уравновешивается силой
трения. Проекция этой силы меняется в зависимости от угла ![]() , поэтому ее необходимо проинтегрировать на интервале
шага. Работа мышц ноги на продвижение
тела равна A = Δh2
, поэтому ее необходимо проинтегрировать на интервале
шага. Работа мышц ноги на продвижение
тела равна A = Δh2 ![]() + Δh2
+ Δh2 ![]() Мощность Р = dA/dt = mgkтр · Δh2 · Lш /Lн .
Здесь принято Lш = Lн = 0,8 м, , Δh1 = 2 Δh 2 . Откуда мощность передвижения для тела
человека массой 80 кг равна 80вт, при kтр =0,05 и скорости 5км/час. Плюс к этому надо добавить мощность на движение самих ног.
То же самое мы получим, если будем рассчитывать потенциальную энергию
колебаний тела на величину
Мощность Р = dA/dt = mgkтр · Δh2 · Lш /Lн .
Здесь принято Lш = Lн = 0,8 м, , Δh1 = 2 Δh 2 . Откуда мощность передвижения для тела
человека массой 80 кг равна 80вт, при kтр =0,05 и скорости 5км/час. Плюс к этому надо добавить мощность на движение самих ног.
То же самое мы получим, если будем рассчитывать потенциальную энергию
колебаний тела на величину ![]() h =
3см (замеренная величина). N = mg
h =
3см (замеренная величина). N = mg
![]() h /
t , где t время половины шага, в течение которого
происходит подъем тела. P = 72 Вт. В работе [13] проведен
расчет энергии шагающего тела с учетом затраты энергии на передвижение ног, но
получены завышенные результаты. По всем
методикам расчета мощность на передвижение равна 70 – 104 Вт, или 84±20%. Она
равна мощности основного метаболизма, который природа свела к минимуму из соображений экономии энергии, которую
животное должно добывать для своего существования. И это относится ко всем
видам биологических тел. Толчком к
исследованию этого вопроса явилась моя собственная мысль, почему на передвижение
по земле биологических тел требуется мощность 1 Вт /кг. Анализ литер.
данных подтвердил этот вывод, все
животные на земле, включая рыб и птиц затрачивают на передвижение в
среднесуточной оценке 1 Вт на кг веса, за исключением животных крайних размеров.
У слона получается величина меньше, 0,8
Вт/кг в спокойном движении, у мышей и
животных меньшей величины расход возрастает, в некоторых случаях до10 – 30
кратного увеличения (как считает Иванов и некоторые другие биологи). Но здесь
они ошиблись, это не энергия на передвижение, это основной метаболизм у мелких
животных возрастает, так как он проходит по необычному пути. Связано это с
увеличением интенсивности кислородного потребления в особых природных
условиях. Но не у всех, пчелы, шмели,
малые птички сохраняют величину 1вт на кг с небольшими отклонениями в обе
стороны. Например, быстрые микробы, у которых просто рассчитать мощность
передвижения Enterobacter,
( длина L=15 мкм, ширина 1мкм, скорость 10 L/сек),
дают удельную мощность 1,2 вт на
кг (собственные измерения и расчеты), импульсная мощность 15 вт/кг. Кокковые микробы имеют малую мощность на передвижение; короткие палочковые, которые вертятся как пропеллер, имеют
большую импульсную мощность, до 20 Вт/кг, но это мощность при избытке
питательной массы, номинальная мощность у них также близка к величине
1Вт/кг±20%. Все проведенные выше
расчеты сделаны для обоснования номинальной мощности на передвижение в
среднесуточной величине.
h /
t , где t время половины шага, в течение которого
происходит подъем тела. P = 72 Вт. В работе [13] проведен
расчет энергии шагающего тела с учетом затраты энергии на передвижение ног, но
получены завышенные результаты. По всем
методикам расчета мощность на передвижение равна 70 – 104 Вт, или 84±20%. Она
равна мощности основного метаболизма, который природа свела к минимуму из соображений экономии энергии, которую
животное должно добывать для своего существования. И это относится ко всем
видам биологических тел. Толчком к
исследованию этого вопроса явилась моя собственная мысль, почему на передвижение
по земле биологических тел требуется мощность 1 Вт /кг. Анализ литер.
данных подтвердил этот вывод, все
животные на земле, включая рыб и птиц затрачивают на передвижение в
среднесуточной оценке 1 Вт на кг веса, за исключением животных крайних размеров.
У слона получается величина меньше, 0,8
Вт/кг в спокойном движении, у мышей и
животных меньшей величины расход возрастает, в некоторых случаях до10 – 30
кратного увеличения (как считает Иванов и некоторые другие биологи). Но здесь
они ошиблись, это не энергия на передвижение, это основной метаболизм у мелких
животных возрастает, так как он проходит по необычному пути. Связано это с
увеличением интенсивности кислородного потребления в особых природных
условиях. Но не у всех, пчелы, шмели,
малые птички сохраняют величину 1вт на кг с небольшими отклонениями в обе
стороны. Например, быстрые микробы, у которых просто рассчитать мощность
передвижения Enterobacter,
( длина L=15 мкм, ширина 1мкм, скорость 10 L/сек),
дают удельную мощность 1,2 вт на
кг (собственные измерения и расчеты), импульсная мощность 15 вт/кг. Кокковые микробы имеют малую мощность на передвижение; короткие палочковые, которые вертятся как пропеллер, имеют
большую импульсную мощность, до 20 Вт/кг, но это мощность при избытке
питательной массы, номинальная мощность у них также близка к величине
1Вт/кг±20%. Все проведенные выше
расчеты сделаны для обоснования номинальной мощности на передвижение в
среднесуточной величине.
На основании проведенного исследования можно
сформулировать энергетический закон природы: расход мощности на
передвижение биологических тел составляет 1 Вт на кг массы тела. Определяется
эта величина всем комплексом биофизических и биохимических процессов в
биологическом теле и является оптимальной величиной для клеточного метаболизма. Шредингер заметил, что
биологические тела состоят из легких элементов, но не понял почему. А причина в экономии энергии даже за счет
собственной массы. Вот почему этот вопрос и требует физико-философского
исследования. Волькенштейн тоже обратил
на это внимание и пришел к выводу, что этот вопрос может быть решен только с
помощью физики – нужен фундаментальный подход, т.е. физический анализ с
математическим обоснованием.
Но борьба биологических тел требует прочности, т.е. в конструкциях тел
надо применять прочные материалы, но прочные материалы, как правило, тяжелые, а
масса должна быть минимальной. Поэтому биологические тела построены из легких
элементов H, C, N, O, P, Ca, Na, K [4].
Кальций, самый тяжелый элемент, но из него строится скелет, именно он
придает организму механическую прочность.
Благодаря легким атомам
плотность молекул низка, а прочность достигается за счет соответствующей конфигурации
самих молекул. Удельная прочность некоторых материалов,
например, оснований крыльев, оказывается выше, чем у железа, титана и др.
тяжелых металлов. Конструкторы
животного мира так располагали атомы в молекулах, чтобы масса тела была
минимальной, прочность максимальной, а средняя плотность тел не превышала
1.1г/см3. И проведенный поиск связи массы с
энергией показал, что масса должна быть минимальной из соображений экономии
энергии, а это значит, что атомы, из которых она состоит, должны быть
легкими.
В заключение возникает вопрос, почему
такая круглая цифра 1 вт, а не 1.5 вт?
Ответ дан при анализе рис 1, 2 и 3. Этот минимум расхода энергии для
биологических тел достигается путем оптимизации всех молекулярно-биохимических
процессов в клетках живых систем.
Поиск оптимального соотношения массы тела, диапазона скорости движения и
требуемой им энергии привел к тому соотношению массы и скорости ее движения в
живом мире, которое построено на экономии энергии. Полученная величина определяется биофизикой и биохимией
клеточного метаболизма, величиной g на
планете, размерами планеты, и от других природных параметров. Оно
определяется возможностью такого построения молекул, которое необходимо для
выполнения всех физических требований борьбы и движения биологических тел в
пространстве их существования, и всем комплексом биохимических превращений,
которые необходимы для нормального существования этих тел в борьбе их за
получение биологической энергии из
других биологических источников в
борьбе видов. Иванов К.В.,
Шмидт-Нильсен и некоторые другие
биологи считают [2,3,8], что в животном мире имеет место большой перерасход
энергии, и что механизмы, созданные
человеком, имеют более высокий КПД. Но
в работах [9,10] доказывается, что
автомобиль имеет КПД 2 %, поэтому
потребляет он 200 – 400 кВт, т.е. на
два порядка большую мощность, чем животные. Но ошибки биологов простительны,
т.к. они не знали этих работ, они появились позднее, когда вопросами основного
метаболизма перестали заниматься, а теорией транспорта еще не начали. И надо
помнить, что главный конструктор биологических тел, сконструировал живые
механизмы намного лучше людей, которые еще очень далеки от понимания сути
устройства этих механизмов.
Энергетически выгоднее делать
шагающие устройства для передвижения, но проще делать транспортные устройства
на колесах.
Все изложенные виды расчета мощности на передвижение по поверхности
планеты дают близкие результаты, так что физический подход к решению
биологических задач оказался оправданным, внесена ясность в определение нормы
расхода энергии при движении на планете. Это также ставит вопрос о пересмотре
принципов построения транспортных средств, имеющих слишком большой расход
энергии по сравнению с животным миром.
Этот перерасход нарушает температурную экологию планеты, особенно в северном
полушарии.
Referat:
Предложено несколько способов определения соответствия массы тела и
требуемой для ее передвижения энергии. Оригинальный физико-философский способ
определения взаимной связи массы тела с необходимой для его движения энергией
представляет интересный ход в научных исследованиях. Применение его в других
отраслях знаний может дать заметное
увеличение скорости научных исследований. Нельзя все время ставить эксперименты
и ждать от них положительных сдвигов в
решении проблемных задач при поиске истины. Пора переходить к логическим
экспериментам, которые позволяют существенно уменьшить объем опытов, отнимающих
много времени и средств. Решена важная
задача поиска принципа построения жизни в природе – это выбор минимума энергии
для определенной массы в заданном диапазоне скоростей. Решение проведено на
принципах физики и механики твердых тел. Дано обоснование необходимости
применения легких элементов в построении биологических тел живой материи по
периодической системе элементов. Работы биологов не позволили им дать ответы на
многие вопросы, так как голое биологическое экспериментирование без
физико-математических расчетов завело их на тупиковую колею исследования
метаболизма животных и связанных с этим вопросов. Образное мышление открывает новый путь исследования научных
проблем.
Таблица основных физических параметров биологических тел
|
Животное |
m кг |
Основной метабол. |
Скорость номинал. |
Скорость максимал |
Мощность номинальн |
Мощность максим.вт |
Удельная мощность |
|
Cлон |
4000 |
3600 вт |
2 км/час |
20км/час |
3500 Вт |
15000 Вт |
0.88 вт/кг |
|
Бык |
800 |
780 |
4 |
40 |
750 |
8000 |
0.9 |
|
Лошадь |
600 |
600 |
4 |
60 |
550 |
3500 |
0.93 |
|
Cтраус |
70 |
|
4,5 |
70 |
70 |
1600 |
1.0 |
|
Человек |
80 |
84 |
4,2 |
30 |
85 Вт |
1200 |
1.06 |
|
Собака |
18 |
24 |
4,3 |
35 |
20 |
110 |
1.1 |
|
мышь |
0.03 |
10 |
0,35 |
3 |
0.03 |
1.6 |
0.86 |
Лит:
1.Kleiber M. The Fire of Life. An
Introduction to Animal Energetics. New
York, Wiley, 454 pp., 1961. 2. Schmidt-Nielsen K. Why is animal
size so impotent? London, New York, пер.260 c. М.М. 1987. 3. Иванов К.П. Основы энергетики организма, Спб.
Наука, т.1 – 5 , 1990 – 2007г. (список лит.) 4. Волькенштейн М.В. Биофизика, М. Наука,
1988.
5. Lehninger A. Biochemistry. N-Y: Worth Publ.
Inc, 1972. 730 p.
6. Гамбарян
П.П.
Бег млекопитающих, Л. Наука,1972. 7. Гинзбург И.Ф. Нерешенные проблемы
фундаментальной физики, Успехи физических наук, т.179, N5, с.525 – 529.
8.
Горшков В.Г. Мощность и скорость передвижения животных разных размеров. Журнал
общей биологии, 1983, т. XLIV,
N5, с.661 – 678, список
лит. 9. Голубев
В.И. КПД транспортных средств, ж.
”Двигатель”, 2006, N2,
с. 44 – 47. 10. Голубев
В.И. Автомобильные проблемы, М. Science Press,
2006. 112c. 11. Кокшайский Н.В Энергетика полета насекомых и птиц, Ж.общей биологии, т.31, N5, с.527–548. 12. Кокшайский Н.В. Изв. АН СССР, МТТ, 1977, N3. 13.
Белецкий В.В. Динамика двуногой ходьбы. Изв. АН СССР, МТТ, 1975, №3,4. 14.
Неймарк Ю.И. Ходьба как управляемые
автоколебания, Изв. АН СССР, МТТ,1983,N4.
15. Casey T.M. A comparison of mechanicl and energetic of flight
cost for hovering sphinx moth, J. Exper.
Biol., 91, 117 – 129, 1981.
16.
Голубев В.И. Функциональная схема двигательной регуляции и стриарного
демпфирования колебаний. Труды РНТО РЭС, т.LV, М. 2000, с.222 – 224.
17. Human Physiology? Edited by Schmidt R.F. and Thews G.,
Springer-Verlag, Berlin-N Y, 1995, v.1,3.
18. Борисов А.В. Биомеханика ходьбы человека, М, 2009г. .
19. Thomas S.P. Metabolism during flight in to species of bats, J. Exper. Biol., 63, 273 – 293,
1975. 20. Wipp B.I. Wasserman
E. Efficiency of muscular work J. appl. Physiol. 1969. V.26, N5, p.644-648 21. Вольтерра М.
Математическая теория борьбы за существование. М. Наука, 1976. 22. Романенко Е.В.
Гидродинамика рыб и дельфинов, М. 2001.