Моцний М. П., Сомова
І. І., Чернетченко[1]Д. В.
Дніпропетровський національний університет ім. Олеся Гончара
ДОСЛІДЖЕННЯ ДІЇ ПОТЕНЦІАЛ-УТВОРЮЮЧИХ ІОНІВ НА
БІОЕЛЕКТРИЧНІ ХАРАКТЕРИСТИКИ РОСЛИН
Метою нашої роботи є
дослідження впливу іонів Са та К на біоелектричні параметри рослин та їх
динаміку. Для спостереження зміни цього стану та подальшого контролю
біоелектричних потенціалів розроблено та застосовано автоматизовану установку
на базі ЕОМ. На основі приведеної комплексної експериментальної установки виконано
обробку відведених потенціалів, під час дії хімічних речовин, що впливають на
зміну концентрації Са та К клітин рослини. Показано відповідність отриманих
експериментальних даних та теоретичних розрахунків на зміненій математичній
моделі Ходжкіна-Хакслі для іонних механізмів Сa, Na та К.
Експериментальна установка.
Експериментальна установка складається із блоку вимірювання «1», який в свою
чергу складається з екранованої камери (об’єкт), в якій розміщені електроди, що
не поляризуються ЕВЛ-1М, рослина, що досліджується, вихідні пристрої
стимуляторів, підсилювач та калібрувальний генератор. Пристрій реєстрації
потенціалів «2» – самописець Endim. Термостимулятор «3», який складається з джерела
живлення (акумулятор із напругою живлення 6 В), реле часу, цифрового регулятора
струму, цифрового вимірювача струму, ключа К3 , інвертора струму
термостимулятора, в основі роботи якого полягає ефект Пельт’є, мідно-ніхромової
вимірювальної термопари. Фотостимулятор «4», який вміщує джерело
живлення (батарея із напругою живлення 4,5 В), цифрового міліамперметра, ключа Кф , блока світлодіодів.
Електричний стимулятор «5», що складається з джерела живлення (батарея із
напругою живлення 9 В, типу «крона»), перемикача Ке та цифрового мікроамперметра. Вивід на
ЕОМ під’єднується до виводів цифрового пристрою – USB-осцилографа, що
зчитує дані та зберігає їх на ЕОМ. Структурна схема такої установки
представлена на рис.1.
Характеристики USB-осцилографа. У якості
пристрою оцифровки даних було обрано USB-осцилограф «IRIS», що забезпечує
наступні режими роботи:
-2-х канальний
осцилограф (маркерні вимірювання, синхронізація, вимірювання напруги і частоти
сигналу, фільтрація).
-2-х канальний
спектр-аналізатор (маркерні виміру, різні віконні функції, фільтрація).
-2-х канальний самописець
(маркерні вимірювання, запис сигналу протягом декількох десятків годин).
Кількість каналів
осцилографа: 2; частота дискретизації: 100 Гц ... 200 кГц; глибина пам'яті:
читання через буфер: 1126 відліків / канал (1 канал), 563 відліків / канал (2
канали) потокове читання: 64K відліків / канал (1 або 2 канали); вхідна
напруга: -20 ... +20 В; розрядність
АЦП: 10 біт; синхронізація абсолютна (по наростаючому / спадаючому фронту),
диференційна (по різниці між сусідніми відліками), зовнішня (по наростаючому / спадаючому фронту ТТЛ рівні);
віконні функції Хаммінга, Ханнінга, Блекман, Блекман-Харріса.
Для поєднання USB-осцилографа ЕОМ було розроблено
універсальний драйвер пристрою. Розрахунки, наступна візуалізація та обробка
оцифрованих даних проводилася за допомогою розробленого програмного
забезпечення на базі візуального середовища програмування LabView.
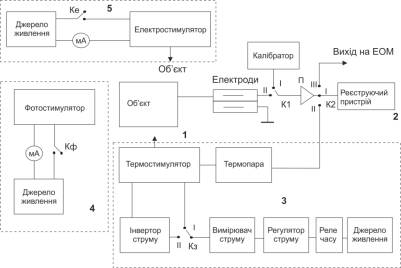
Рис.1.
Блок-схема експериментальної установки
Робота установки. Режим «калібрування».
Перемикач К1 встановлено в положенні І, К2 у положенні І, відповідно. З виходу калібратора на вхід
підсилювача подається калібрувальний синусоїдальний сигнал, який записується на
самописець.
Режим
«термостимуляція». Перемикач К1 – у положенні ІІ, К2 – І. Лист рослини
розташовано в екранованій камері. Розміщуємо на листи контакти
термостимулятора, які зроблені в вигляді защіпки. Над контактом стимулятора
розміщуємо активний електрод, для відведення електричних потенціалів, а
пасивний – на не стимульованій поверхні листа. В положенні «охолодження» К3
встановлено в положенні І. Параметри імпульсу подразнення, формуються реле часу
та регулятором струму. Для позиції «нагрів» необхідно встановити перемикач К3
в положення ІІ. Для вимірювання температури використовують режим «термопара». В
цьому режимі сплав термопари розташовують між контактами термостимулятора, а
перемикач К2 – переводять в положення ІІ. В цьому режимі
пристрій реєстрації записує динаміку
зміни температури на контактах термостимулятора. ЕРС термопари обрано 56
мкВ/град 0С.
Режим
«фотостимуляція». Перемикач К1 – у положенні ІІ, Ке – у
положенні І. Лист рослини розміщують над світлодіодом, який генерує певну
довжину хвилі світла. Світлодіод має спеціальний інфрачервоний фільтр для
зменшення нагріву листа. Активний електрод розташовують на світовій площині
листа, а пасивний – на неосвітленій частині.
Освітленість
регулювалася зміною струму світлодіода, який вимірюється цифровим
міліамперметром. Освітленість вимірювалась за допомогою фотометру. Тривалість
стимулу задавалася за допомогою реле часу, або в ручному режимі, при тривалій
стимуляції.
Режим
«електростимуляція». Перемикач К1 розташовано у положенні ІІ, К2
– І. Електричний стимул подається на електроди відводу, за допомогою реле, що
по’єднане з реле часу, яке
формує тривалість стимулу. Струм регулюється реостатом та вимірюється цифровим
мікроамперметром. При увімкненні стимулятора, реле відключає електроди від
підсилювача, вхід підсилювача «закорочується», а електроди з’єднує з
електростимулятором. При закінченні стимулу подразника, реле вмикає на вхід
підсилювача електроди, та «разкорочує» його входи. Таким чином струм
подразнення не подається на вхід підсилювача.
Вихід
на ЕОМ. Перемикач К2 – в положенні ІІІ. При цьому вихід підсилювача
з’єднується входом цифрового USB-осцилографу, що під’єднується до відповідного
порту ЕОМ.
Клітинне моделювання. . Використовувався
математичний пакет MATLAB - для
обробки та аналізу масиву отриманих даних. Було розроблено математичну модель
нейрону із здатністю генерації активних електричних потенціалів на основі
механізму Ходжкіна-Хакслі. Параметри, що характеризують характер генерації
обчислювався динамічно в процесі експерименту, за допомогою застосування
апроксимації вхідних даних динаміки електричного потенціалу. Модель із
параметрами описувалася за формулою (1).
![]() (1)
(1)
де Еi –
потенциал рівноваги i-го іонного каналу, gi – електрична провідність каналів, що визначається за формулами (2):
![]()
![]() (2)
(2)
![]()
Функції φ(t) та x(t) описують швидкість реакції каналів та
визначаються звичайно за методом підтримки постійного потенціалу на мембрані
клітини, при цьому диференційні рівняння (2) дають розв’язки (3).
![]() , (3)
, (3)
![]()
З експериментальних
даних ми обчислювали сталі часу активації та інактивації каналів рослинних
клітин τφ та τx,
відповідно.
Результати
та їх обговорення
Дослідження та обробка отриманих потенціалів
рослин було проведено на базі розробленого програмного забезпечення. Зміну
потенціалу із часом у цифровому форматі зчитуємо через USB-порт ПК або
портативного ПК за допомогою USB пристрою (осцилограф IRIS), відбувається
автоматичне зберігання отриманих цифрових даних зміни потенціалу із часом у
форматі *.dat. У вікні клієнтської програми відбувається аналіз отриманих
даних, і запис головних показників у базі даних програмного забезпечення а
саме: амплітуди піків
холодових (або теплових) потенціалів, із відповідними часовими відмітками;
процентні співвідношення амплітуд піків відведених потенціалів рослини, що
характеризують динаміку протікання процесу; поточні налаштування часових та
амплітудних параметрів для калі бровки сигналу, та кількісної оцінки отриманих
сигналів та візуалізації шкал графічної залежності «потенціал – час».
Повторюючи експеримент необхідну кількість разів
отримуємо результати із заданою точністю. Паралельно із цифровим записом
сигналу потенціали також фіксувались за допомогою самописцю, для контролю та
перевірки. Для листів кукурудзи були отримані наступні результати вимірювань.
Запис одиночного
потенціалу показано на рис. 2, на якому можна бачити стандартну електричну
відповідь на холодовий подразник («холодовий потенціал»), без застосування згладжую
чого фільтра. Увімкнення та
налаштування фільтра, що відрізає мережеву перешкоду, а також випадкові
високочастотні перешкоди у мережі, дає
змогу отримати більш чіткий сигнал, що можна спостерігати на рис. 3 (а та б).
Після налаштування фільтру було проведено запис електричний відповідей від
електродів с поверхні рослини необхідну кількість разів, для отримання заданої
точності вимірів, що в даному випадку склала 15 дослідів.
а б
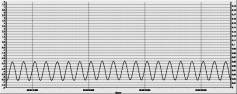

Рис.2. Запис стандартного сигналу калібровки від генератор (а). Холодовий потенціал без
додаткової фільтрації сигналу (б).
а б
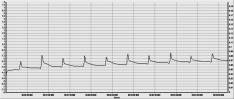

Рис.3. Холодові потенціали рослини для (а) постійної
стимуляції та одиничний холодовий потенціал (б) за умови застосування
фільтруючих засобів.
В результаті обробки отриманих
потенціалів, ми отримали графік залежності амплітуди потенціалів від інтервалу
часу між першим і наступним подразниками, при цьому ми бачимо, що з часом
співвідношення А,% значно зростало, з поступовою стабілізацією на рівні 130%
відносно першої відповіді на подразник, як показано на рис. 4.
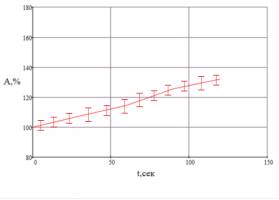
Рис.4. Графік
залежності амплітуди електричних відповідей рослини, відносно амплітуди першого
піка потенціалу із часом.
Що свідчить про процес
активації внутрішньоклітинних механізмів адаптаптації рослини до подразники, та
поступовий процес звикання до стимулів, без зростання амплітуди відповіді на
нього.
Для отриманих даних залежності
потенціалу від часу, були розраховані часові параметри та провідності іонних
каналів активних клітин за формулами (2), g1 = 32.455 та g2 =
110.231 , що досить близько до значень провідності каналів натрію та калію gNa та gK . Що досить достовірно
підтверджує походження зареєстрованих потенціалів. Періодичні осцилятивні
відповідні, що спостерігалися на рис.3а, є активною електричною відповіддю
клітини на подразних – потенціал дії, у основі якого лежить взаємодія Na+
- K+
апарату клітини. Деяка похибка електричних провідностей від табличних значень
спостерігається за рахунок наявності у клітині струмів інших типів, а також
пасивного струму витоку іонів з клітини.
Висновки
Головним результатом
роботи слід вважати розробку повністю автоматизованої системи обробки
експериментальних даних для реєстрації електричних потенціалів рослин, яка
представляє собою комплексну програмно-апаратну установку для вимірів. Була
виконана перевірка відповідності результатів отриманих за допомогою
автоматичної обробки та обробки даних з самописця. Результати свідчать про
досить високий процент точності результатів отриманих за цифровими даними, а
похибка, що накопляється за рахунок обмеженості вимірювального пристрою, а
також зовнішніх перешкод, як то мережева перешкода 50 Гц, знаходиться в
допустимих межах, та не впливає суттєво на результат. Додаткову роль в цьому
відіграє програмно реалізований фільтр, із змінними параметрами та
згладжувальний фільтр, який в даній роботі було обрано типу фільтра Баттерворта
3-го порядку, що дозволяє отримати якісні дані для подальшої обробки.
Було розроблено
математичну модель, що описувала процеси генерації електричних імпульсів у
клітині. В результаті розрахунку параметрів моделі, ми встановили, що в основі
електричної відповіді лежить селективна зміна електричної провідності мембрани
клітини для різних типів іонів, для цього випадку було встановлено, що це іони Na+ та К+.
В свою чергу це є досить достовірним підтвердженням походження електричних
відповідей рослини на подразник. Надпорогові потенціали відповіді були
потенціалами відповіді, що досить точно описуються нашою моделлю. Отже, ми
вставили зв’язок із потенціалами активності, що реєструвалися з поверхні
рослини та внутрішньоклітинними процесами, що в ній протікають та розроблена
математична модель збудливої клітини може бути застосована для опису цих
процесів.
Література
1. Вплив термічної стимуляції на біопотенціали листя кукурудзи / М. П. Моцний, Н.П.Боцьва, О.В.Єліна, С.В.Власова, І.П.Матвій // Вісник Дніпропет-ровського університету. Біологія. Медицина. – 2010. – Т.2. – С. 65–70.
2. Евдокимов Ю. К. LabView для радиоинженера: от виртуальной модели до реального прибора / Ю. К. Евдокимов, В. Р. Линдваль, Г. И. Щербаков. – М.: ДМК Пресс, 2007. – 399 с.
3. Пятыгин С. С. Сопряжение генерации потенциала действия в клетках растений с метаболизмом: современное состояние проблемы / С. С. Пятыгин, В. А. Воденеев, В. А. Опритов // Усп. соврем. биол. – 2005. – Т. 125. – С. 534–542.