Биологические науки/10.
Генетика и цитология
Медицина/8. Морфология
Антонова Е.И., Шпак О.Ю., Сагалбаева Л.Е.
Омский государственный педагогический университет, Россия
Гистотопография, цитотипы печени животных с различной системой
терморегуляции и стратегией метаболической адаптации
Регенерация печени, а
следовательно и поддержание тканевого гомеостаза реализуется благодаря межклеточным
стромально-паренхимным цитокоммуникациям. Сложность структуры,
полифункциональность, быстрота вовлечения печени в деструктивные и репаративные
процессы - все это определяет неослабевающий интерес исследователей к проблемам
ее регенерации: источники, масштабы, механизмы. Высокая интенсивность
метаболических процессов у эндотермных животных (птицы) и развитие в эволюции у
эктотермных животных (амфибии) механизмов метаболической «депрессии»,
становление у них новых механизмов устойчивости к гипоксии, реоксигенации
способствовали усложнению, экологической специализации и совершенствованию
морфологии печени, которая, по сути, является «периферическим интегратором
метаболизма» [3, 6]. В связи с этим цель нашей работы является изучить
отличительные особенности и вектор формирования в эволюции клеточного
микроокружения, как основу тканевого гомеостаза печени животных с различной
системой терморегуляции, стратегией метаболической и экологической адаптации.
Исследования проводили
на двух видах животных - амфибии вида Rana terrestris Andrzejewski, 1832 (Rana arvalis Nilsson,
1842), и
птицы вида Columba livia (forma domestica) Gmelin, 1789. Эксперимент поставлен на 30 половозрелых особях самцах
(амфибии трехлетки и голуби шести месяцев постнатального развития). Для
гистологических исследований образцы печени фиксировали в 10%-нейтральном
формалине, заливали в парафин. Стромально-паренхимное соотношение клеточных
типов печени, пролиферативную активность и полиплоидизацию гепатоцитов
определяли с помощью выявления антител к белкам-маркерам этих клеток [4]. Для
выявления процессов пролиферации и полиплоидизации парафиновые срезы окрашивали
антителами к PCNA гепатоцитов (Ig – мышиные МАТ; клон РС
10; разведение 1:100; DAKO), клетки Ито антителами
к десмину стрептавидин-биотиновым методом (Ig – МАТ; клон – десмин D33;
разведение 1:30; DAKO, DENMARK). Антигены выявлялись
после предварительной демаскировки методом HIAR (Нeat-Induced Antigen Retrieval).
После инкубации с первичными антителами инкубировали биотинилированными вторыми
антителами (Link, DAKO LSAB+Kit
Peroxidase), со стрептавидином, коньюгированным с
пероксидазой хрена (Streptavidin, DAKO
LSAB+Kit Peroxidase).
В качестве субстрата пероксидазной реакции использовали раствор АЭК и перекиси
водорода. Гистохимическая реакция определения активности эндогенной пероксидазы
на парафиновых срезах проводилась с целью выявления клеток Купфера по методу
МакФи [5].
Статистическую обработку
полученного материала осуществляли с помощью пакета прикладных программ ”STATISTICA-6”.
За критический уровень значимости принимали р=0,001. Выявляли критерий согласия
χ2 Пирсона, равенство дисперсий σ (Фишера-Снедекора),
согласно которым определяли возможность применения параметрических и
непараметрических методов анализа статистических данных (критерий Стьюдента, критерий
однородности Вилкоксона-U).
В ходе исследований выявлено, что гистологическое
строение печеночного ацинуса в сравнительном ряду интактных животных изменяется
от трубчатого типа организации у амфибий, к трубчато-трабекулярному у птиц. От
экто- к эндотермным животным увеличивается количественная плотность печеночных
ацинусов, что свидетельствует об интенсификации процессов метаболизма и
становления в эволюции полифункциональности органа. В печени обоих групп
животных определяется большое количество меланомакрофагальных центров, в
формировании которых участвуют клетки Купфера. Главная роль таких центров заключается в депонировании эндогенных и экзогенных веществ с
дальнейшей деструкцией, детоксикации и повторном их использовании [2].
Анализ показателей пролиферации/полиплоидизации
(двуядерные формы гепатоцитов) и полиплоидизации (PCNA-позитивные гепатоциты)
выявил, что у амфибий распределение двуядерные формы гепатоцитов носит венуло-портальный
характер. Процент двуядерных гепатоцитов от общего количества гепатоцитов
составляет 11,6% (рис. 1). У птиц распределение двуядерных гепатоцитов носит
порто-венулярный характер, а общее количество двуядерных гепатоцитов составляет
9,7% (рис. 2).

Рис. 1. Распределение PCNA – позитивных и
двуядерных гепатоцитов в условиях физиологической нормы у амфибий вида Rana terrestris в пределах печеночного ацинуса (примечание:
*р=0,001).
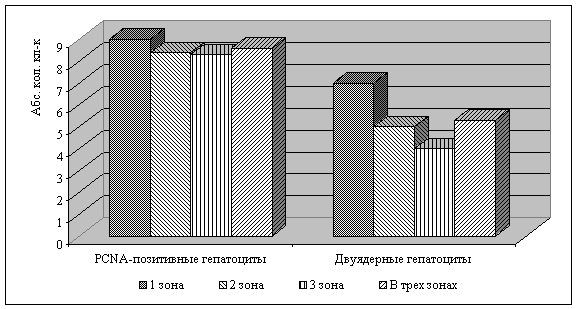
Рис. 2. Распределение PCNA – позитивных и
двуядерных гепатоцитов в условиях физиологической нормы у птиц вида Columba livia в пределах
печеночного ацинуса (примечание: *р=0,001).
Показатель полиплоидизации - количество PCNA-позитивных
гепатоцитов, выше у птиц и во всех зонах ацинуса. Тогда как у амфибий максимальное
количество PCNA-позитивных гепатоцитов выявлено в области портального
тракта (рис. 1, 2). Следовательно, у эктотермных животных ведущей формой
регенерации гепатоцитов является пролиферация гепатоцитов за счет
ацитокинетических митозов. У эндотермных животных ведущей формой является
процесс полиплоидизации.
Процесс регенерации гепатоцитов регулируется как
аутокринно, так и парактинными цитокоммуникативными механизмами [1]. Так на
фоне большей активности ацитокинетических митозов у амфибий выявлено самое
большое количество пероксидазо-позитивных клеток Купфера (рис. 3). При этом у амфибий распределение данного типа клеток в пределах ацинуса носит
равномерный характер, у птиц отмечается преобладание органоспецифичных
макрофагов в области портального тракта.
Наибольшее количество десмин-позитивных клеток
Ито выявлено у птиц, при этом выявляются видовые особенности топографии данной
популяции стромальных клеток у изучаемых видов животных в пределах зон
печеночного ацинуса (рис. 4).
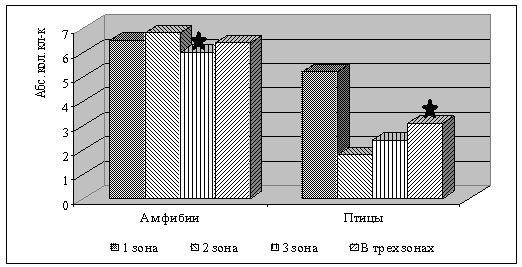
Рис. 3. Количество пероксидазо
- позитивных клеток Купфера в условиях физиологической нормы у амфибий вида Rana terrestris и у птиц вида Columba livia в пределах печеночного ацинуса (примечание: *р=0,001).
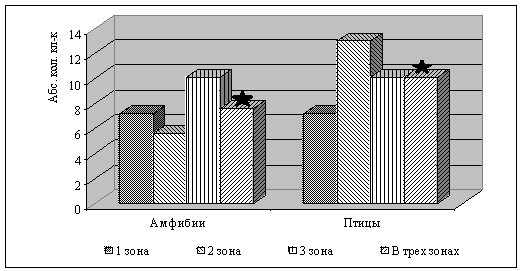
Рис. 4. Количество десмин-позитивных клеток Ито в условиях физиологической
нормы у амфибий вида Rana terrestris и у птиц вида Columba livia в пределах печеночного ацинуса (примечание: *р=0,001).
Полученные нами данные позволяют
сделать заключение о том, что в процессе эволюции у животных с различной
системой терморегуляции клеточный состав печени не менялся; наибольшее количество
органоспецифичных макрофагов определяется у эктотермных, а клеток Ито у
эндотермных животных; показатели пролиферации гепатоцитов по зонам ацинуса
носят разнонаправленный характер, при этом ведущей формой регенерации
гепатоцитов у птиц является полиплоидизация, у амфибий
пролиферация/полиплоидизация. Выявленные количественные
отличия стромально/паренхимных соотношений, за счет синтеза цитокинов,
обеспечивают адаптивные сдвиги гомеостаза, для нужд локального региона
организма – печени, а так же проявляют зависимость со стратегиями метаболизма
изучаемых групп животных: амфибии – толерантная, птицы - резистентная.
Литература:
1. Антонова Е.И.
Реактивность и пластичность тканевых компонентов печени в сравнительном ряду
позвоночных в норме и после гипертермии [Текст]: автореф. дис….док. биол. наук:
03.00.25 /Е.И. Антонова. – А., 2009.- 31с.
2. Антонова, Е.И.
Ультраструктурные проявления первичной компенсаторно-приспособительной реакции
гепатоцитов животных с различной системой терморегуляции после воздействия
гипертермии / Е.И. Антонова // Морфология.- 2008.- №4.- 24-28с.
3. Калашникова, М.М. Особенности ультраструктуры клеток печени
в сравнительно-морфологическом ряду животных я их значение [Текст] / М. М. Калашникова
// Бюлл. экспер. биол.- 1996. - Т. 121, №
6. - 604-609с.
4.
Киясов, А.П. Современные технологии морфологических
исследований [Текст] / А.П. Киясов.- Казань: КГМУ.-2000.-38с.
5.
Маянский,
Д.Н. Новые рубежи гепатологии [Текст] / Д. Н. Маянский, Э. Виссе, К. Декер. -
Новосибирск, 1992. - 264с.
6. Scholz,
H. Adaptational responses to hypoxia [Тext] / H. Scholz // Am. J. Physiol.
Regul. Integr. Comp. Physiol.- 2002.- Vol. 282.- P. 1541-1543.