Биологические науки/9. Биохимия и биофизика.
К.б.н. Ловягина Е.Р, Чуркина Л.А., Школьников Д., к.б.н.
Солнцев М.К., д.б.н. Семин Б.К.
Московский государственный университет
им.М.В.Ломоносова, Россия
Пути окисления экзогенных доноров в
фотосистеме 2,
не содержащей марганец
В фотосинтезирующих организмах
пигмент-белковый комплекс фотосистемы 2 (ФС2) осуществляет окисление воды и
выделение кислорода. Этот процесс является одной из важных биоэнергетических
реакций, поскольку представляет собой основной источник кислорода в атмосфере.
Вследствие этого исследование фотосинтетического окисления воды имеет большое
фундаментальное и технологическое значение.
Окисление
воды в ФС2 осуществляется каталитическим центром – кислород выделяющим
комплексом (КВК). Благодаря исследованиям последних лет стали известны многие
особенности строения ФС2 и структуры КВК. Функционирование каталитического
центра обеспечивает работа электрон-транспортной цепи. После поглощения кванта
света вторичный донор электронов тирозин-161 белка D1 (YZ) передает
электрон фотоокисленному первичному донору электронов
Р680+ и, в свою очередь, окисляет Mn-кластер КВК. Источником электронов
для работы КВК служат молекулы воды. В нативной ФС2
окислитель YZ• недоступен
для компонентов внешней среды и может окислять только КВК. При экстракции КВК
(кластера Mn4-Ca и трех периферических белков, закрывающих этот
кластер) из ФС2 [ФС2(-Mn)] YZ• становится
доступным для экзогенных доноров электронов и окисляет их с высокой
эффективностью. Однако, помимо YZ•-зависимого
участка окисления в ФС2(-Mn) появляется еще один участок окисления экзогенных доноров
(низкоаффинный). Природа окислителя на этом участке неизвестна. Существуют экспериментальные указания на то,
что роль альтернативного окислителя может играть His. Логично предположить, что это – D1-His190 [Tommos & Babcock, 2000; Semin et al., 2005],
расположенный вблизи YZ на
одинаковом с ним расстоянии от Р680. Моделирование поверхности ФС2 без КВК
показывает, что His190
расположен фактически на поверхности белковой глобулы.
Такое положение в отсутствие КВК делает его доступным для экзогенных доноров
электронов, таких как Mn(II), Fe(II), дифенилкарбазид (ДФК), гидроксиламин.
Ранее было показано, что основной путь транспорта электронов через YZ может быть заблокирован катионами Fe [Semin et
al., 1995; 2002]. В отсутствие Mn они
эффективно связываются с нативными Mn-связывающими участками КВК и экранируют YZ от экзогенных доноров. В таких
блокированных препаратах функционирует только один путь транспорта электрона –
от низкоаффинного участка окисления к Р680+,
что очень удобно для его изучения.
Информативным
методом, используемым для исследования работы электрон-транспортной цепи ФС2,
является метод термолюминесценции [Vass & Govindjee, 1996; Култышева и др.,
2001]. При удалении марганцевого кластера на кривых высвечивания
термолюминесценции появляется специфическая полоса при –18oС, так называемая «АТ-полоса».
С применением модификаторов гистидина и карбоксильных групп установлено, что
эта полоса обусловлена рекомбинацией зарядов восстановленного первичного
акцептора Q-A и окисленного редокс-активного
гистидина [Ono & Inoue, 1991].
В
задачу настоящей работы входило выяснение участия остатка гистидина-190 белка D1 реакционного центра ФС2 в альтернативном переносе электрона
от искусственного донора ДФК к Р680 с помощью регистрации АТ-полосы
термолюминесценции.
В
результате проведенной работы установлено, что блокирование основного пути
транспорта электронов через YZ катионами Fe в препаратах ФС2(-Mn) сопровождается ~50% подавлением АТ-полосы термолюминесценции.
Такой эффект, вероятно, обусловлен тем, что связывание катионов Fe с высокоаффинным участком
(участком высокого сродства к Mn) приводит к затруднению индукции His+.
Модификатор
гистидиновых остатков диэтилпирокарбонат
(ДЕПК) в концентрации 20 мМ и выше полностью
подавляет АТ-полосу в препаратах ФС2 без Mn. При этом
электронный транспорт от искусственного донора электронов ДФК через
альтернативный окислитель сохраняется на 60%. Результаты с ДЕПК, на первый
взгляд, указывают на то, что His не
принимает участия в альтернативном транспорте. Уменьшение скорости электронного
транспорта в модифицированных катионами Fe препаратах ФС2(-Mn) на 40%, видимо, носит неспецифический характер, поскольку
такое же уменьшение наблюдается в контрольных экспериментах при обработке ДЕПК неблокированных железом препаратов ФС2(-Mn). Однако
не исключено, что при связывании ДЕПК с His-190 (вероятным альтернативным окислителем), расположенным на
расстоянии водородной связи от YZ,
происходят пространственные изменения, приводящие к восстановлению доступности YZ для ДФК. В этом случае в
поставленном эксперименте мы регистрировали
электронный транспорт не через альтернативный окислитель, а через YZ.
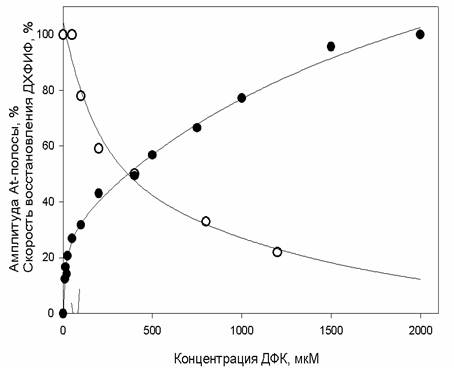 В
последующих экспериментах было показано, что увеличение концентрации
искусственного донора электронов ДФК приводит к увеличению скорости
электронного транспорта, что в свою очередь количественно коррелирует
с уменьшением амплитуды полосы АТ в препаратах ФС2 без Mn с
блокированным с помощью катионов железа YZ (рис.1).
В
последующих экспериментах было показано, что увеличение концентрации
искусственного донора электронов ДФК приводит к увеличению скорости
электронного транспорта, что в свою очередь количественно коррелирует
с уменьшением амплитуды полосы АТ в препаратах ФС2 без Mn с
блокированным с помощью катионов железа YZ (рис.1).
Рис. 1. Зависимость амплитуды АТ-полосы (o) и скорости транспорта электронов
ДФК-Р680-ДХФИФ (·) от
концентрации ДФК в препаратах ФС2(-Mn) с блокированным YZ.
Такая
корреляция между увеличением скорости электронного транспорта по альтернативному
пути и уменьшением амплитуды АТ-полосы при
увеличении концентрации ДФК указывает на участие His-190 в альтернативном переносе электрона.
Полученные
результаты в целом свидетельствуют в пользу участия His-190 в альтернативном переносе электрона от экзогенных
доноров к Р680.
Литература:
1.
Култышева М.Ю.,
Ловягина Е.Р., Кузнецов А.М., Солнцев М.К., Семин
Б.К., Иванов И.И. (2001) Сравнительное исследование влияния экзогенных
доноров электронов фотосистемы II на АТ-полосу термолюминесценции. Биохимия, 66, 884-890.
2. Ono T., Inoue Y. (1991) Biochemical evidence for histidine
oxidation in photosystem II depleted of the Mn-cluster for O2-evolution. FEBS Lett., 278, 183-186.
3. Semin B. K., Ivanov I. I., Rubin
A. B., Parak F. (1995). High-specific binding of Fe (II) at the Mn-binding
site in Mn-depleted PSII membranes from spinach. FEBS
Lett., 375,
223-226.
4. Semin B. K., Ghirardi
M. L., Seibert M.
(2002). Blocking of electron donation by Mn
(II) to Y (Z*) following incubation of Mn-depleted photosystem II membranes with Fe (II) in the light.
Biochemistry, 41 (18), 5854-5864.
5. Semin B. K., Lovyagina E. R., Timofeev K. N., Ivanov I. I., Rubin A.B., Seibert M. (2005). Iron-Blocking the High-Affinity Mn-Binding Site in Photosystem II
Facilitates Identification of the Type of Hydrogen Bond Participation in
Photon-Coupled Electron Transport via YZ+. Biochemistry, 44 (28), 9746-9757.
6. Tommos C., Babcock G. T. (2000). Proton and hydrogen currents in fotosynthetic water oxidation. Biochim.
Biophys. Acta, 1458,
199-219.
7. Vass I., Govindjee (1996). Thermoluminescence
from photosynthetic apparatus. Photosynthesis Res. 48, 117-126.