С.А. Онищук
Кубанский
государственный университет, Россия
ЭФФЕКТИВНЫЕ УСКОРЕНИЯ ДИНАМИКИ БИОСИСТЕМ
При неизменных внешних
условиях в частично равновесной открытой системе в стационарном состоянии,
близком к термодинамическому равновесию, скорость прироста энтропии за счет
внутренних необратимых процессов достигает отличного от нуля постоянного
минимального положительного значения. Этот принцип минимума прироста энтропии,
или теорема Пригожина, представляет собой количественный критерий для
определения общего направления самопроизвольных изменений в открытой системе
вблизи равновесия. Если система находится в стационарном состоянии, то она не
может самопроизвольно выйти из него за счет внутренних необратимых изменений.
При отклонении от стационарного состояния в системе должны произойти внутренние
процессы, возвращающие ее к стационарному состоянию, что соответствует принципу
Ле-Шателье - устойчивости равновесных состояний. Поэтому любое отклонение от
устойчивого состояния вызовет увеличение скорости продуцирования энтропии.
Однако ранее не
обсуждался вопрос, каким именно способом система придет в стационарное
состояние. В данной работе исследуется динамика показателей системы при их изменении
в случае выхода ее из равновесного состояния за счет внешних воздействий и
возвращении в равновесное состояние.
Вопросами динамики
биологических процессов активно занимается биокинетика [1]. В ней в частности
рассматриваются функциональные зависимости показателей биосистемы при переходах
из равновесного состояния в неравновесное и обратно. Причем если в первом
случае выявлено много различных вариантов формул для разного вида систем, то во
втором практически всегда зависимость экспоненциальная или близкая к ней.
Для биологических систем
ранее было установлено, что выведение различных веществ, например, лекарств из
организма происходит с линейной скоростью или близкой к ней, а концентрация его
снижается по экспоненте. Была установлена линейная зависимость скорости от
концентрации, а поскольку скорость изменения концентрации является
дифференциалом от самой концентрации, то при интегрировании получаем
экспоненту.
Но дифференциалом от
скорости является ускорение, а этот вопрос не рассматривался. Не был также
затронут вопрос об оптимальном ускорении изменения показателей биосистемы из
равновесного состояния в неравновесное при возмущающем воздействии.
В данной работе
предлагаются следующие постулаты.
1. Постоянное ускорение
изменения показателей любой системы, биологической (или небиологической),
является оптимальным для динамических процессов системы. Это в первую очередь
относится к переходу биосистемы из равновесного состояния в неравновесное
2. При переходе
биосистемы в равновесное состояние ускорение изменения значений показателей пропорционально
скорости.
Согласно первому
постулату скорость, как интеграл ускорения, должна возрастать линейно, а сам
показатель, как интеграл скорости – по параболе. Подобным образом в работе [2] использовали
квадратичные зависимости для моделирования относительного потока клеток в периферическую
кровь при анемии. Второй постулат предполагает линейную зависимость скорости от
показателя, и экспоненциальную зависимость показателя от времени. Примером
является диссоциация лиганд-рецепторных комплексов [1].
При одновременном
прохождении процессов как нарастания, так и спада показателя, обе зависимости
перемножаются, и динамика показателя изменяется по S-функции [2].
Данное утверждение было
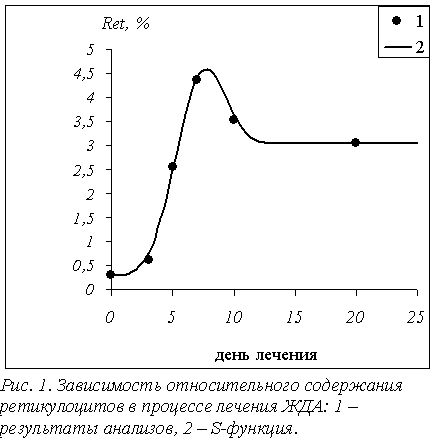
проверено на примере динамики показателей
крови при лечении анемий различного генеза. В частности, динамика относительного количества ретикулоцитов при лечении железодефицитной анемии в точности соответствовала обоим постулатам (рис. 1) с
некоторыми незначительными отклонениями вследствие особенностей кроветворительной
системы каждого конкретного человека. Более того, по S-функции
изменялись все показатели крови.
 Если первый постулат относится к обоим процессам, то имеет
место принцип, который можно было бы назвать принципом постоянных ускорений. Любая
система при выведении ее из состояния равновесия изменяется и возвращается к состоянию
равновесия при постоянных ускорениях изменения показателей.
Если первый постулат относится к обоим процессам, то имеет
место принцип, который можно было бы назвать принципом постоянных ускорений. Любая
система при выведении ее из состояния равновесия изменяется и возвращается к состоянию
равновесия при постоянных ускорениях изменения показателей.
Литература:
1.
Варфоломеев С.Д.,
Гуревич К.Г. Биокинетика. Практический курс. М.: ФАИР-ПРЕСС, 1999. 720 с.
2.
Барановская И.Б., Онищук
С.А. Моделирование динамики показателей крови при лечении анемий различного
генеза. Врач и информационные технологии. 2008. №2. С.48-54.
3.
Свищенко В.В., Стахин
Н.А. Математическое моделирование
компенсации анемии дополнительным потоком клеток в периферическую кровь, возникающим
вследствие анемии. Бюллетень сибирской
медицины, 2008. № 4, С. 44-47.