Биологические науки/ 8.Физиология человека и
животных.
Д.м.н. Мустафин А. Г.
Российский
национальный исследовательский медицинский университет им. Н. И. Пирогова.
Москва.
Околочасовые ритмы интенсивности синтеза РНК и белков в популяциях
нервных клеток разных отделов нервной
нервной системы крыс и их взаимоотношения с суточными ритмами транскрипции и
трансляции.
Аннотация Установлено, что в изученных отделах нервной системы крыс отчетливо
можно выделить две группы популяций нейронов с характерным временным
расположением акрофаз активности хроматина.
Активность генома невроцитов осуществляется импульсно и периодически.
Формирование суточного ритма активности генетического и белоксинтезирующего
аппаратов нейронов происходит за счет вхождения отдельных групп клеток в
процессы, обеспечивающие транскрипцию и трансляцию, в колебательном режиме.
Ключевые
слова: суточные и ультрадианные ритмы, синтез РНК и белков, популяции нервных
клеток, разные отделы нервной системы крыс.
I. Введение В пределах одной функциональной системы имеют место
ритмы различных частотных диапазонов, что, по-видимому, способствует строгой
иерархической организации разнообразных процессов внутри организма.
Ультрадианные ритмы отражают особенности протекания основных физиологических
процессов, могут характеризовать механизмы краткосрочных физиологических
реакций организма животных и осуществляться на разных уровнях их организации [1]. Современные молекулярные исследования
свидетельствуют, что возбуждение посредством вторичных и третичных посредников
распространяется вглубь цитоплазмы и ядра нервной клетки и оказывает влияние на
процессы синтеза белков и РНК. Экспрессируемые геномом нейронов генные продукты
включаются в системные реакции различного качества, определяя молекулярную
интеграцию деятельности нейрона [2]. Основываясь на собственных и литературных данных В.Я.
Бродский [3] характеризует выявленные ритмы с периодом около 1 ч (околочасовые
ритмы) количественных изменений белков как фундаментальное общеклеточное
свойство. Ультрадианные ритмы связаны с цикличностью метаболических процессов
и, тем самым, являются эндогенными. Многообразие ритмических процессов,
протекающих в биологических объектах и согласованных во времени между собой и с
изменяющимися условиями внешней среды рассматривается как временная организация
живых систем [4, 5].
II. Постановка задачи. Целью проведенного нами
исследования являлось изучение ультрадианных ритмов включения меченного уридина
и лейцина в суммарные РНК и белки клеток разных отделов нервной системы крыс.
III. Результаты. Эксперимент поставлен на 350 крысах-самцах линии Wistar массой около 170-210 г, содержавшихся в стандартных
условиях при фоторежиме 12:12 часов (свет с 8 до 20 час). Животных (по 3-4 на
каждую временную точку исследования) забивали с 15-минутными интервалами на
протяжении суток. Для изучения интенсивности транскрипции и трансляции брали
спинномозговые и краниальные шейные симпатические ганглии; поясничное утолщение
спинного мозга; кору мозжечка; супрахиазматические ядра гипоталамуса;
зрительную и сенсомоторную области коры большого мозга. Срезы указанных органов
культивировали in vitro. Часть срезов каждого органа инкубировали с 3Н-уридином,
вторую половину - с 3Н-лейцином. Интенсивность синтеза суммарных РНК
и белков регистрировали методом жидкостной сцинтилляционной авторадиографии.
Рассчитывали абсолютное и относительное включение изотопов в суммарные РНК и
белки. Параметры активности синтеза первичных генных продуктов изучали на
протяжении 22 ч с 0800 до 0600 ч. Полученные данные были обработаны методом
спектрального анализа с использованием пакета программ обработки временных
последовательностей.
На рис.1 представлены данные по изучению
суточной динамики синтеза суммарных РНК и белков срезами спинномозговых узлов
крыс.
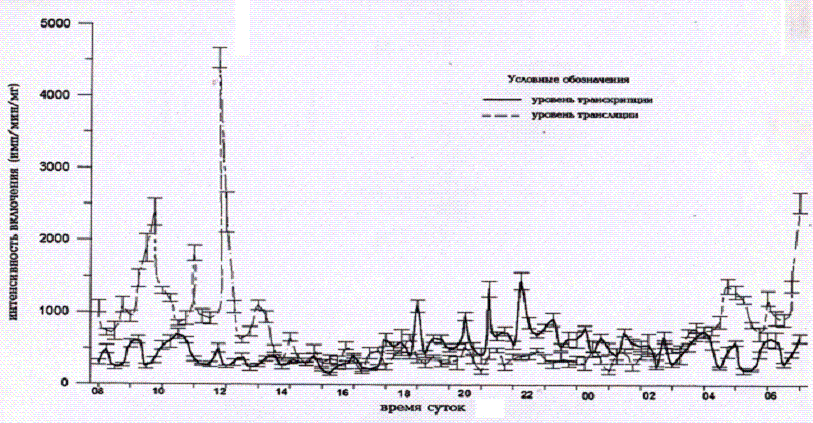
Рис.1. Изменения
интенсивности включения Н3-уридина и Н3—лейцина клетками
спинномозгового узла на протяжении суток при
забое животных через 15-минутные интервалы.
Максимальная интенсивность включения 3Н-уридина
приходится на 21 ч 15 мин, а 3Н-лейцина - на 11 ч 45 мин,
минимальные - в начале и в конце светового периода соответственно (P<0,01). С помощью программы для выявления скрытых
периодичностей обнаружены колебания матричной активности хроматина с периодами
11,5 ч (амплитуда равна 23,4%), 2,5 ч (амплитуда равна 8,9%), и 55 мин
(амплитуда равна 3,6%). Колебания активности белоксинтетического аппарата
составили 12,5, 3,5 ч и 79 мин (амплитуды равны 33,9, 12,7 и 1,9% соответственно).
Сопоставление суточного ритма и периодов ультрадианных ритмов клеток
спинномозговых узлов крыс показало, что в активной фазе суточного ритма
активности генетического аппарата период околочасовой составляющей был равен
39,5±3,6, а в пассивной
фазе суточного ритма - 56,0±4,2 мин. (P<0,02). В то же время период
околочасовой составляющей активности белоксинтетического аппарата был равен в
активную фазу ритма 70,7±6,8, а в пассивную 42,0±5,7 мин (P<0,01). Околочасовая
составляющая в активную фазу суточных ритмов синтеза первичных генных продуктов
отличается длительностью периода от пассивной фазы ритма. Наличие подобных гармоник
показывает, что активность генома осуществляется импульсно и периодически. Так
же можно высказать предположение о том, что при формировании суточного ритма
активности генетического и белоксинтезирующего аппаратов отдельные группы
клеток разных отделов нервной системы крыс входят в процессы обеспечивающие
транскрипцию и трансляцию в колебательном режиме. Это согласуется с данными
ряда авторов, показавших ритмичность активности клеток ряда органов [6,7].
Для срезов краниальных шейных
симпатических ганглиев, поясничного утолщения спинного мозга; коры мозжечка;
супрахиазматических ядер гипоталамуса так же как и в спинномозговых узлах
максимальная интенсивность включения 3Н-уридина приходится на начало
темнового периода суток, а 3Н-лейцина - на конец темнового и начало
светового периода суток. Обнаружены так же колебания матричной активности
хроматина с периодами 11-12,0 ч (амплитуда равна 10-13,5%), 2,6-3,5 ч
(амплитуда равна 18,9%), и 56-48 мин (амплитуда равна 2,8-5,6%). Периодики
интенсивности включения 3Н-лейцина составили 11,6-12,9, 2,9-3,7 ч и
37-53 мин (амплитуды равны 25,6-32,9, 11,5-13,7 и 1,5-3,2% соответственно).
Максимальные значения относительного
включения 3Н-уридина срезами соматосенсорной коры (рис.2) отмечены в
начале светового периода суток (0800-1200 ч).
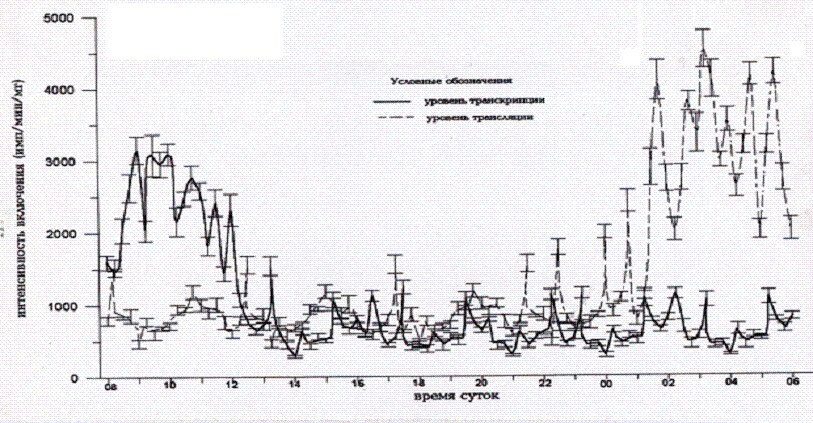
Рис.2. Изменения
интенсивности включения Н3-уридина и Н3—лейцина клетками
соматосенсорной коры большого мозга на протяжении суток при забое животных через 15-минутные интервалы.
Минимальные значения обнаружены в остальные
исследованные часы суток (Р<0,01). Ультрадианные периодики составили 9,2,
3,7 ч и 44 мин (амплитуды - 16,8, 9,7 и 2,3% соответственно). Ритмическая
активность функционирования белоксинтезирующего аппарата характеризуется
достижением максимальных значений ближе к середине темнового периода суток
(0130-0600 ч, Р<0,01). Показаны так же 11,2, 4,3 часовые и 52 минутные периодики (амплитуды - 18,9,
10,2 и 1,8% соответственно). В активной фазе суточного ритма матричной
активности хроматина наименьший период ультрадианной составляющей (49,3 мин)
был на 22% выше, чем в пассивной фазе суточного ритма (38,3 мин.). В темный
период суток минимальный период ультрадианной составляющей интенсивности
трансляции был равен 48,5 мин, а в дневные часы суток - 54,4 мин. Суточная динамика активности генома клеток срезов
зрительной коры сходна с таковой, для соматосенсорной коры головного мозга
крыс.
IV. Выводы. Таки образом, в изученных отделах нервной
системы крыс отчетливо можно выделить две группы популяций нейронов с
характерным временным расположением акрофаз активности хроматина. Биоритмологические характеристики ритмов активности
хроматина имеют свои особенности в клеточных системах, отличающихся
функционально и по положению: максимальные уровни экспрессии генов клеток
спинномозговых узлов супрахиазматических ядер гипоталямуса, краниальных шейных
симпатических ганглиев, коры мозжечка и спинного мозга соответствуют по срокам
повышенному уровню активности крыс (ночные часы суток); максимальные уровни
экспрессии генов нейронов зрительной и соматосенсорной областей коры большого
мозга соответствуют по времени дневной рецепции света. Активность генома
невроцитов осуществляется импульсно и периодически, формирование суточного ритма
активности генетического и белоксинтезирующего аппаратов нейронов происходит за
счет вхождения отдельных групп клеток в процессы, обеспечивающие транскрипцию и
трансляцию, в колебательном режиме;
Литература.
1. Биологические ритмы. Т. 2 / Под ред. Ю. Ашоффа. – Москва:
Мир, 1984. – 414 с.1
2. Судаков
К.В. Интегративная деятельность нейрона: специфика и пластичность системных
механизмов.//.Журн. высш. нервн. деятельности. 1993. Т.43. вып.2 с. 289-300.
3. Бродский В.Я., Нечаева Н.В. Ритм синтеза белка.-
М.: Наука, 1988,- 240с
4. Романов Ю.А., Междисциплинарный характер
исследований временной организации биологических систем и их значение для
медицины. // Биология и медицина / Под ред. Ю.А. Овчинникова. - М., 1985. - с. 90-103
5. Koseska A.,
Ullner E., Volkov E., Kurths J., Garcia-Ojalvo J. Cooperative differentiation
through clustering in multicellular populations // J. Theor. Biol. — 2010. — Vol. 263.
— P. 189–202.
6 Рыбаков
В.П. Клеточно-популяционные закономерности механизма формирования суточного ритма
репродукции клеток. Автореф дисс. докт. мед. наук. М.1991
7. Бродский, В. Я.; Дубовая, Н. Д.; Звездина, Т. К.;
Фатеева, В. И.; Мальченко, Л. А. Дофамин дезорганизует ритм синтеза белка,
нарушая самоорганизацию гепатоцитов in vitro. Бюллетень экспериментальной
биологии и медицины. - 2013. - Т. 156, № 7. - С. 48-50. -