Ямборак Виталий Васильевич, аспирант кафедры теоретической кибернетики
Киевского Национального университета имени Тараса Шевченка.
Компьютерное моделирование
когнитивных
процессов в мозге человека
В последнее время все больше работ
посвящено моделированию работы головного мозга, в частности процессов мышления.
В связи с большой сложностью объекта исследования, является чрезвычайно
непростым построение концепций, которые не противоречат многочисленному
экспериментальному материалу и согласуются с известными психологическими и
психофизическими феноменами. Одной из немногочисленных таких концепций является
концептуальная модель, предложенная в [1]. Именно она берется за основу.
Разработан
компьютерный инструментарий для построения и имитационного моделирования работы
биологических нейронных сетей на клеточном
уровне (уровне нейронов и синапсов). Инструментарий рассчитан на работу с нейронными сетями
размеров порядка ![]() нейронов и
представляет собой компьютерную программу, входными данными которой является
нейронная сеть, а также входы самой сети, описанные на XML. Разработан XML-язык
для представления биологической нейронной сети как множества нейронов и
синапсов. Разработанный инструментарий был использован для
построения и тестирования модели обонятельной луковицы. Проведенные
эксперименты подтвердили несколько гипотез относительно концептуальной модели.
нейронов и
представляет собой компьютерную программу, входными данными которой является
нейронная сеть, а также входы самой сети, описанные на XML. Разработан XML-язык
для представления биологической нейронной сети как множества нейронов и
синапсов. Разработанный инструментарий был использован для
построения и тестирования модели обонятельной луковицы. Проведенные
эксперименты подтвердили несколько гипотез относительно концептуальной модели.
Описание каждого эксперимента состоит
из двух пунктов:
1. Экспериментирование. Описание проделанных действий
экспериментаторами. На этом этапе проводилось построение нейронной сети для
инструментария и подача требуемых входов на эту сеть, а также снятие выходных
данных сети.
2. Интерпретация результатов
экспериментирования. В
этом пункте делался акцент на том, каким образом полученные результаты согласуются
с концептуальной моделью.
Далее детально описан каждый эксперимент. Отметим, что
планирование и проведение экспериментов совпали, что связано с отсутствием сложностей, которые могли возникнуть во
время экспериментирования.
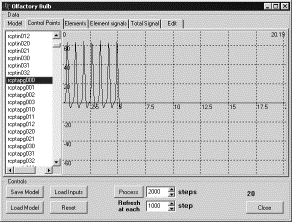
Выходные
сигналы и опознание. На
входы рецепторного нейрона, соответствующего одной обонятельной зоне [2],
подавался входной сигнал, достаточный для активации, на отрезке времени от 0 до
5. Выход снимался с соответствующей обонятельной зоне митральной клетки. В
результате рецепторным нейроном на интервале времени, на котором подавались
входные сигналы, генерировались потенциалы действия (ПД), потом они
прекращались (рис. 1). Но генерация ПД продолжалась в митральной клетке и после
временной отметки 5 (рис. 2). Также как и в митральной клетке, после
прекращения подачи входного сигнала на вход рецепторного нейрона, продолжалась
генерация ПД и в кисточковой клетке (рис. 3), что может быть обусловлено
большим весом связи митральной и кисточковой клетки.
В
данном эксперименте анализировались выходные сигналы с митральной клетки, а не
с кисточковой, в отличие от того, как это делалось в [2]. Данная
несогласованность с [2] вызвана тем, что в концептуальной модели придерживаются
позиции, которая частично отличается от той, которая имеется ввиду в работах [2,
3].
Поскольку
митральная клетка возбуждались после того, как вход на рецепторный нейрон был
прекращен, то мы получали вторичный всплеск [1]. Это соответствует опознанию
входного стимула, так как возбуждалась митральная клетка, которая соответствует
возбужденному рецепторному нейрону. Факт того, что ПД генерировались после
окончания подачи входных стимулов на модель, также свидетельствует о
кратковременном запоминании стимула.
|
|
|
|
Рис. 1. Генерация ПД в рецепторном нейроне. |
Рис. 2. Генерация ПД в митральной клетке. |
|
|
|
Рис. 3.
Генерация ПД в кисточковой клетке |
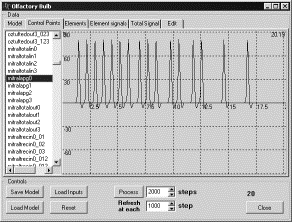
Проверка
существования «умственного взгляда». Входной сигнал
подавался на постсинаптические мембраны кисточковых клеток (а не на рецепторные
нейроны) на отрезке времени от 0 до 5. После этого подача входного сигнала
прекращалась. На временном интервале от 0 до 5 генерировался ПД в кисточковой
клетке. После временной отметки 5 еще некоторое время происходила генерация ПД,
потом она прекращалась (рис. 4). С началом подачи сигнала на вход кисточковой
клетки начинается также генерация ПД в митральных клетках. После прекращения
подачи сигнала на вход кисточковой клетки в момент времени 5 генерация ПД в
митральных клетках еще продолжалась (рис. 5).
В
этом эксперименте происходило возбуждение митральной клетки, т.е. возбуждались
квазирецепторные нейроны. В концептуальной модели это соответствует
«представлению» объекта в тот момент, когда во внешней среде он не представлен.
Другими словами, было подтверждено существование «умственного взора» [1] в
обонятельной луковице.
|
|
|
|
Рис. 4. Генерация ПД в кисточковой клетке |
Рис. 5. Генерация ПД в митральной клетке |
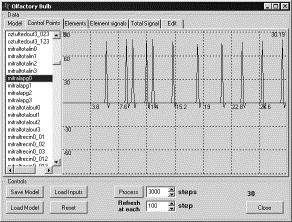
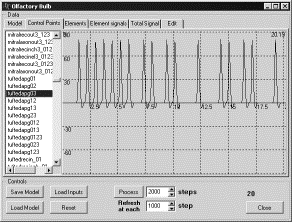
Кратковременная
память как прямая и обратная связь кисточковых и митральных клеток. В данном
эксперименте были отключены связи от кисточковых клеток к митральным, которые
проходят через зернистые клетки [2]. При экспериментировании на
обонятельной луковице была необходимость в некотором уточнении, поскольку связи
между кисточковым и митральными клетками в данном случае не непосредственные, а
осуществляются через зернистые клетки [2], и гораздо сложнее, чем
простейший случай в концептуальной модели. Как следствие реализаций может быть
несколько.
|
|
|
Рис. 6. Генерация ПД в рецепторном нейроне |
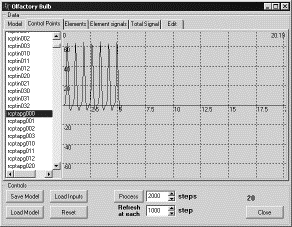
Таким
образом, в нейронной сети, моделирующей обонятельную луковицу, отключались
связи от кисточковых к митральным клеткам тремя способами:
1. путем
удаления зернистой клетки со всеми связями, входящими и выходящими из нее;
2. путем
удаления всех связей от кисточковых клеток к зернистым и от зернистых к митральным;
3. путем
удаления всех связей от зернистых клеток к митральным.
Во
всех трех случаях подавался сигнал на входы рецепторных нейронов на интервале
времени от 0 до 5. В результате рецепторные нейроны до момента времени 5
генерировали ПД, после этого момента ПД, естественно, прекращались (рис. 6).
|
|
|
|
а) |
б) |
|
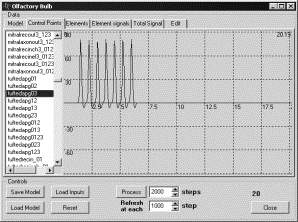
Рис. 7. Генерация ПД в кисточковой клетке а) при
удалении связей 1 и 2 способами; б)
при удалении связей 3 способом. |
|
|
|
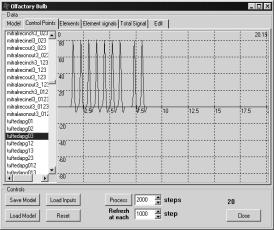
|
Рис. 8 Генерация
ПД в митральной клетке при удалении связей во всех трех случаях |
Активность
основных нейронов (митральных и кисточковых клеток) несколько отличалась для
разных способов реализации эксперимента.
Рассмотрим
реализацию первым и вторым типами. При подаче сигнала на входы рецепторного
нейрона в кисточковых и митральных клетках генерировался ПД. После прекращения
подачи входного сигнала генерация ПД в них сразу же прекращается (рис. 7а, 7б,
8). При удалении связей способом 3 после прекращения подачи сигнала на входы
рецепторных нейронов кисточковые клетки генерирует дополнительный ПД в ответ на
свой же входной сигнал, который поступил на них через зернистые клетки.
Указанный
эксперимент подтверждает известную гипотезу о том, что замкнутые нейронные
циклы выполняют роль кратковременной памяти. Таким образом, повторных всплесков
в митральных клетках не происходило при разрыве циклических связей, указанных
выше. Это и подтверждает роль циклической связи как реализации механизмов
кратковременной памяти.
Обучение путем синаптической пластичности.
При подаче входных сигналов на рецепторные нейроны с моделированием наращивания
весов синаптических связей реализуется один из механизмов долговременной и
кратковременной памяти – долговременная и кратковременная синаптическая
пластичность соответственно.
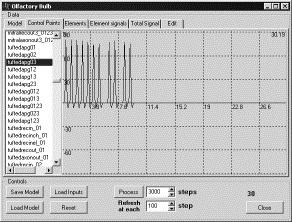
На
интервалах времени от 0 до 5 и от 10 до 15 единиц на рецепторные нейроны
подавался входной сигнал. Выходные сигналы снимались с кисточковых и митральных
клеток. В рецепторных нейронах на
соответствующих интервалах времени генерировались ПД (рис. 9).
|
|
|
Рис. 9 Генерация
ПД в рецепторных нейронах на интервалах времени от 0 до 5 и от 10 до 15 |
При
подаче входных стимулов на протяжении интервала времени от 0 до 5 генерация ПД
митральными и кисточковыми клетками длилась дольше, чем подавался входной
сигнал на рецепторные нейроны (рис. 9-11).
При
подаче адекватных входных стимулов, между которыми небольшой интервал (5
временных единиц), при работе инструментария было заметно, что на втором
интервале подачи входных стимулов (от 10 до 15) выходной стимул длился гораздо
дольше, чем при первой подаче входов на интервале (от 0 до 5) (рис. 9-11).
|
|
|
|
Рис. 10. Генерация ПД в кисточковых
клетках на интервале времени от 0 до 15 |
Рис. 11. Генерация ПД в митральных
клетках на интервале времени от 0 до 15 |
Синаптическая
пластичность соответствует в концептуальной модели усилению весов прямых и
обратных связей, т.е. реализован один из механизмов обучение в понятиях
концептуальной модели. Поскольку при увеличении весов синапсов в сложных
нейронных взаимодействиях кисточковых, митральных и зернистых клеток выходы
митральных и кисточковых клеток продолжались дольше, то можно утверждать, что
проявление синаптической пластичности полностью согласуется с предполагаемой ее
ролью в концептуальной модели.
[1] З.Л.Рабинович.
О естественных механизмах мышления и интеллектуализация ЭВМ. // Кибернетика и
системный анализ, 2003, №5, с.82-88.
[2] Г.С.Воронков,
В.А.Изотов. Компьютерное моделирование механизмов обработки информации в
обонятельной системе. I. Модель структурно-функциональной организации
нейронных элементов обонятельной луковицы и рецепторного эпителия // Биофизика.
2001, том 46. вып.4, с.696-703
[3] Г.С.Воронков,
В.А.Изотов. Компьютерное моделирование механизмов обработки информации в
обонятельной системе. II. Механизмы опознания и кратковременного
запоминания в обонятельной луковице: результаты компьютерного
экспериментирования // Биофизика. 2001, том 46, вып. 4, с.704-708.