Биологические
науки/ 2. Структурная ботаника и биохимия растений
Д.
б. н., проф. Злобин Ю.А.
Сумский национальный аграрный университет, Украина
ИНДИВИДУАЛЬНОЕ
И ПОПУЛЯЦИОННОЕ РЕАГИРОВАНИЕ РАСТЕНИЙ НА СТРЕССОВЫЕ ФАКТОРЫ
Благодаря неподвижности, растительные организмы более часто подвержены разного рода экологическим стрессам по сравнению с животными, которые имеют возможность активно избегать неблагоприятных условий существования (миграции, кочевки, выбор мест гнездования и т.п.). Локальные эколого-ценотические условия произрастания оказываются, таким образом, важным регулирующим фактором, действующим как на отдельные особи растений, так и на их популяции в целом.
Стресс у растений возникает под действием обширного комплекса
эколого-ценотических неблагоприятных воздействий, и его проявления также
оказываются комплексными и разноуровневыми. Рецепторами восприятия стрессовых
факторов как первичные дискретные единицы живой материи являются отдельные
особи растений - индивидуумы. Их реагирова-ние на них
отличается значительной сложностью. Как общее правило, практически любой эколого-фитоценотический стресс ведет к подавлению
продукционного процесса, и общий размер фитомассы
растения снижается. В итоге происходит общая миниатюризация особей растений,
при которой пропорционально уменьшению размера общей фитомассы уменьшается
размер всех частей растения. Но такое явление не частое. Его можно наблюдать
главным образом в посевах культурных растений в случае неоптимального загущения
стояния культурных растений. Так, при анализе посева Medicago falcata c нормой высева 6, 8, 12 и
Более типична и характерна в первую очередь для растений природных экосистем компенсаторная адаптация к стрессу, при которой уменьшение общего размера фитомассы особи у отдельных ее структурных частей вызы-вает: а) пропорциональное уменьшение отдельных размерных показателей, б) стабильное сохранение их размера или в) увеличение отдельных размерных морфопараметров. Такой внутренней перестройкой растения реагируют на стрессовую обстановку, а в некоторых случаях и приспосабли-ваются к ней. Число вариантов подобных трансформаций достаточно велико, так как зависит от жизненной формы растений и реализуемой им стратегии жизни. Биологическая интерпретация таких трансформаций не простая, поскольку у растений наблюдаются как адаптивная, так и стрессовая пластичность [4]. Первая повышает приспосабливаемость растения к неблагоприятным условиям, вторая отражает уровень ингибирования физиолого-биохимических и формообразовательных процессов при стрессе.
Фактически ростовые и формообразовательные стрессовые изменения составляют единый комплекс, в котором имеют место и чисто стрессовая и адаптационная компоненты. Общеизвестен «синдром избегания затенения» у
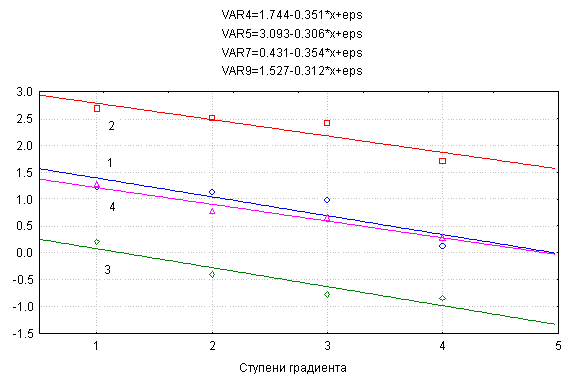
Рис. 1.
Уравнения регрессии и график изменения четырех морфопара-метров особей растений
Medicago sativa
по градиенту возрастания плотности стояния растений в посеве. 1 - надземная
фитомасса, г, 2 - размер листовой поверхности, см2,
3 - фитомасса соцветий, г, 4 - число соцветий, шт. Значения морфометрических
параметров даны в форме логарифмических преобразований.
растений [5]. Он включает в себя изменение направления аллокации веществ, удлинение стебля, уменьшение разветвленности и изменение уровня репродукции. На рис. 2 представлены результаты изменения четырех морфопара-метров Alliaria petiolata по градиенту нарастания эколого-фитоценотического стресса [3]. Как уравнения регрессии, так и их визуализация свидетельствуют, что параллельность линий регрессии отсутствует: направления трансформации значений морфопараметров в этом случае индивидуализированы, отражая разные их тренды. При снижении размера надземной фитомассы особей A. petiolata в ответ на возрастающий стресс в еще большей степени снижается фитомасса генеративных структур, что указывает на падение уровня аллокации веществ в цветки и плоды. В меньшем масштабе, но тоже
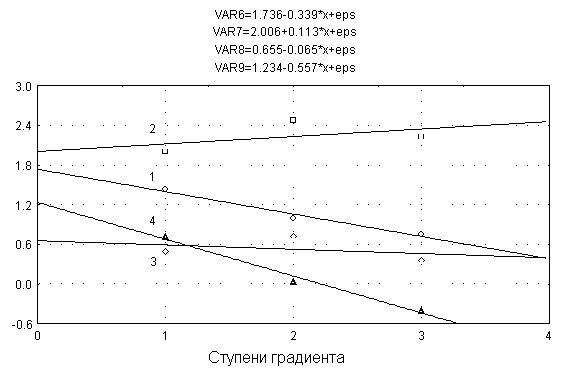
Рис. 2.
Уравнения регрессии и график изменения четырех морфопара-метров особей растений
Alliaria petiolata
по градиенту нарастания эко-лого-фитоценотического стресса. 1 - надземная
фитомасса, г, 2 - размер листовой поверхности, см2,
3 - количество боковых ветвей, шт., 4 - фи-томасса генеративных структур, г.
Значения морфометрических пара-метров даны в форме логарифмических
преобразований.
снижается разветвленность главного побега. Это, безусловно, стрессовые компоненты. Как компенсирующая адаптация у растений одновременно существенно возрастает размер листовой поверхности, что, очевидно, обеспечивает выживание растений, но не сохранение уровня репродуктивного давления особей на экотоп.
В целом, регрессионный анализ дает возможность выявлять масштабы изменения роста и формообразования растений при стрессе и даже идентифицировать чисто стрессовые и адаптационные перестройки в формообразовании особей в условиях стресса.
Для всех природных экосистем характерна выраженная микромозаич-ность местообитаний. Она ведет к тому, что практически в любом местооби-тании уровень воздействия стресса на разные особи оказывается неодинако-вым. Соответственно неодинаковы и изменения продукционного процесса, роста и формообразования у таких растений. Возникает внутрипопуляционная неоднородность особей растений. Ее иногда описывают термином поливариантность онтогенеза [1], а как общебиологическое явление она получает объяснение в концепции виталитета растений [2]. Виталитет особей рассматривается как отражение их общей жизнеспособности, выраженной в росто-вых и структурных параметрах растения, а виталитет популяции показывает соотношение в ней особей растений разного виталитета (процветающих, промежуточных и угнетенных).
Стрессовое ингибирование особей растений в популяциях может оце-ниваться по-разному: а) по одному наиболее характерному признаку, в качестве которого часто выступает «размер» особи, понимаемый либо как общая ее фитомасса, либо как надземная фитомасса, либо как высота и т.п., б) по набору из трех ключевых для уровня жизнедеятельности растения при-знаков или в) по достаточно полному их набору, включающему как статисти-ческие метрические, так и динамические метрические и аллометрические признаки [2]. В любом случае оцениваемые таким способом особи растений в локальной популяции при их репрезентативной выборке составляют статистический ряд распределения, в котором уровень стрессового подавления особи возрастает справа налево.
Компьютерное моделирование и обширный полевой материал, в т.ч. и полученный разными исследователями, показывает, что разнообразие особей в популяции может быть сведено к пяти основным моделям (рис. 3). В местообитаниях, близких к оптимальным с мало выраженной микромозаичностью распределение особей в репрезентативной выборке соответствует типу А. В этом случае в популяциях преобладают особи со средними морфометрическими показателями, особей с отклонениями от этих показателей, чем сильнее эти отклонения, тем меньше. Такой тип часто свойственен эдификаторам. Тип А хорошо аппроксимируется нормальным статистическим распределе-нием. Тип Б характерен для растений, произрастающих в местообитаниях с выраженной мозаичностью и примерно равной площадью основных элементов мозаичности. Встречаемость особей разной размерности здесь примерно одинакова. Тип Б аппроксимируется либо нормальным распределением с низкими значениями эксцесса, либо равномерным статистическим распределением. Тип В характерен для растений контраст-ных местообитаний с наличием элементов мозаики, благоприятных для растений данного вида, и элементов, крайне неблагоприятных. На этой основе появляются двухвершинные типы распределения. Типы Г и Д свойственны популяциям растений, произрастающим соответственно в благоприятной и неблагоприятной среде. Распределение особей в статистических рядах в этих случаях асимметрично. В виталитетном анализе
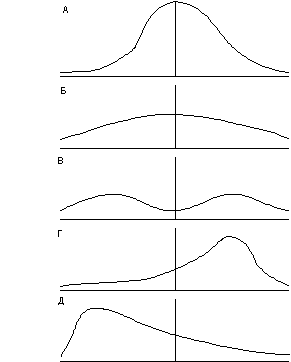
Рис. 3. Основные типы распределения особей растений по их размеру в популяциях. А, Б, В, Г, Д - основные типы распределения особей в популяциях растений по уровню их стрессового ингибирования.
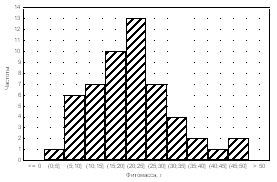
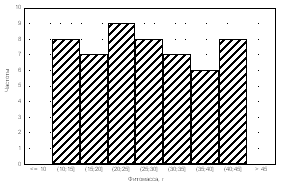
Medicago
falcata Alopecurus pratensis
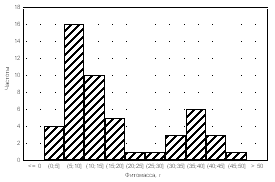
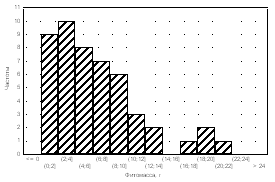
Plantago
major
Amaranthus retroflexus
Рис. 4. Частоты распределения фитомассы особей растений в популяциях.
они соответствуют процветающим и депрессивным популяциям, тогда как типы А, Б и В оцениваются как равновесные популяции.
Частотный анализ распределения особей в популяциях достаточно информативен. Так, при анализе посевов Medicago falcata (рис. 4) было установ-
лено, что тип А распределения особой по размеру надземной фитомассы в
популяции был получен только для варианта норма высева 8 кг/га при междурядьях
В целом, анализ структуры популяций с установлением характера и степени страсс-трансформации отдельных особей и их доли в популяциях дает богатый информационный материал и характеризует как состояние популяций, так и экосистемы, в которых они произрастают.
Литература
1. Жукова Л.А. Поливариантность онтогенеза луговых растений // В сб. «Жизненные формы в экологии и систематике растений». - М.: МГПИ, 1986. - С. 104-114.
2. Злобин Ю.А. Теория и практика оценки виталитетного состава це-нопопуляций растений // Ботан. журн., 1989. - Т. 74, № 6. - С. 769-781.
3.
Злобин Ю.А. Структурная интеграция особей растений // Nauka: teo-ria i praktika - 2007. - Przemyśl,
2007. - Tym 4. -
S. 37-41.
4. Ackerly D.,
№ 11. - P. 979-995.
5. Sultan S. Phenotypic plasticity in plants: a case study in ecological
deve-lopment // Evol. a. Developm., 2003. - Vol. 5, №
1. - P. 25-33.