Арефьев Ю.Ф.1,
Мамедов М.М.2
1Доктор биол. наук, профессор кафедры экологии, защиты леса и лесного
охотоведения, 2Кандидат биол. наук,
ассистент кафедры экологии, защиты леса и лесного охотоведения
Воронежская государственная
лесотехническая академия
ЭКОЛОГО-ГЕНЕТИЧЕСКИЙ КОНТРОЛЬ
ПАТОГЕНЕЗА
В ЛЕСНЫХ ЭКОСИСТЕМАХ
Аннотация
Развивается
принцип контроля патогенеза в лесных экосистемах, основанный на интеграции
экологических и генетических механизмов защиты насаждений. Лесные экосистемы
рассматриваются с позиций концепции детерминированного хаоса. Методика
исследований включала комплекс натурных фитопатологических обследований лесных
насаждений и лабораторных анализов состояния популяций патогенных грибов. На
примере мучнистой росы дуба и корневой губки доказана эффективность инбридинга
в подавлении популяций патогенов. Предложены
схема лесных насаждений, способствующих развитию микоризы и ограничению миграции
спор патогенных для древесных пород грибов. Рекомендован переход от
формирования наиболее распространённых в настоящее время гомогенных лесных насаждений
к формированию устойчивых к инфекционным болезням гетерогенных лесных
экосистем.
Ключевые слова: экология, генетика, контроль,
патогенез, экосистема, интеграция, инфекция
I. Введение
Патогенез, понимаемый как возникновение и развитие инфекционных
болезней, в лесных экосистемах специфичен. Специфика в том, что патогенные
грибы способны размножаться как сексуально, так и асексуально, в то время как
их хозяинные древесные растения в природе размножаются только сексуально, то
есть со сменой фаз ядра.
В результате в
популяциях патогенных грибов наиболее вирулентные штаммы доминируют, но в
популяциях патогенных грибов наиболее
микорезистентные биотипы не могут закрепить свою повышенную
резистентность в потомстве. Второе биологическое преимущество патогенных грибов
заключается в том, что для них характерен быстрый темп смены генераций, в то
время как у лесных древесных растений смена генераций происходит значительно медленнее.
В результате патогенные грибы более успешны в борьбе за существование, чем их
хозяинные древесные породы.
Цель работы заключалась в развитии принципа контроля
патогенеза на основе интеграции экологических и генетических механизмов защиты лесных
насаждений от патогенных организмов. Ранее этот аспект дискутировался в работах
исследователей в разных странах [1 – 3].
Методика
исследований включала комплекс натурных фитопатологических обследований лесных
насаждений в Среднерусской лесостепи и лабораторных работ. В качестве
тест-объектов были наиболее массовые патогенные грибы – мучнистая роса дуба (Erisiphae alphitoides) и корневая губка (Heterobasidion annosum).
В результате исследований была изучена многофакториальная зависимость
патосостояния насаждений. В качестве регулирующих механизмов наибольшее
значение имели инбридинг в популяциях патогенных организмов, естественный
отбор, конкуренция, искусственный отбор, микоризообразование, мутуализм
деревьев.
Прикладной аспект работы в том, что предложены в зависимости
от условий произрастания два типа (с групповым и линейным размещением деревьев,
мозаичность насаждений) формирования
биорезистентных лесных экосистем.
II.
Постановка проблемы
Проблема
контроля патогенеза в лесных экосистемах заключается в том, что для большинства
современных искусственно созданных лесных насаждений характерен высокий уровень
хаотичности. Хаотичность в данном случае понимается как апериодичность детерминированного поведения динамичной
биологической системы, чувствительной к
начальным условиям её состояния. Относительно небольшие изменения в окружающей
среде или внутрипопуляционном состоянии могут привести к значительным
изменениям всей системы. Задача контроля хаотической системой заключается в
снижении степени хаотичности биологической системы, в повышении предсказуемости
динамики биосистемы.
Как биологические
системы, лесные насаждения характеризуются динамичностью и многофакторностью,
для них характерна высокая чувствительность к изменениям в окружающей среде и
внутрипопуляционным изменениям. Задача заключается в том, чтобы хаотическое
поведение конкретной системы преобразовать в регулируемое. При этом
долгосрочные прогнозы становятся менее зависимыми от неконтролируемых
воздействий.
Исследования
проводились в насаждениях Среднерусской лесостепи. Жизнеспособность (V) деревьев и насаждений оценивалась по 5-ти балльной
шкале (Табл. 1). Оценка жизнеспособности деревьев (насаждений) определялась по
формуле:
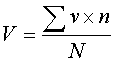 ,
( 1 ) где V –
средневзвешенный балл жизнеспособности,
v – баллы жизнеспособности
отдельных деревьев, n – число
деревьев конкретного балла, N – общее
число учтённых деревьев.
,
( 1 ) где V –
средневзвешенный балл жизнеспособности,
v – баллы жизнеспособности
отдельных деревьев, n – число
деревьев конкретного балла, N – общее
число учтённых деревьев.
Таблица 1: Шкала оценки
жизнеспособности (V) деревьев
|
Состояние
жизнеспособности |
Баллы |
|
Здоровые деревья – без признаков
ослабления |
5 |
|
Ослабленные деревья – кроны слабо
ажурные |
4 |
|
Больные деревья – кроны
значительно ажурные |
3 |
|
Отмирающие деревья – кроны отмерли на 75-80 % |
2 |
|
Отмершие деревья – без признаков
жизни |
1 |
Развитие
болезней определялось по формуле:
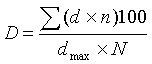 , (
2 )
, (
2 )
где D – развитие
болезни, %, d – поражённость
растения, dmax –
максимальный балл применённой шкалы, n – число растений конкретного балла, N – общее число учтённых растений.
III.
Результаты
Особенности развития
мучнистой росы дуба (Erysiphae alphitoides Griffon et Maubl.) в условиях экологической изоляции
на достаточно малой площади и в условиях открытых насаждений представлены в
таблице 2.
Таблица 2: Особенности развития
мучнистой росы в открытых и экологически изолированных
насаждениях дуба черешчатого
|
Условия
произрастания |
Развитие болезни, D % |
Размеры конидий, µм |
Параметры
клейстотеций |
||
|
Длина |
Ширина |
Диаметр, µм |
Плотность размещения, n/cм2 |
||
|
Открытые насаждения дуба |
78 |
32 |
18 |
99 |
64 |
|
Изолированные на ≈ 0,25 га участки
дуба |
16 |
24 |
13 |
74 |
8 |
Примечание. 1. Участки дуба площадью ≈ 0,25 га были изолированы
насаждениями сосны и берёзы. 2. n – число клейстотеций.
Как следует из таблицы 2, ключевые
параметры популяционного развития мучнистой росы в экологически изолированных
на относительно малых участках дубовых насаждений значительно ниже, чем в
открытых насаждениях дуба. Этот факт свидетельствует о снижении
приспособленности (fitness) патогена, то есть его выживаемости
в условиях его репродуктивной изоляции на относительно малых участках
произрастания хозяинной древесной породы. Снижение приспособленности патогена
является следствием инбридинга – значительного повышения вероятности
идентичности двух аллелей в потомстве
патогена. При этом возрастает гомозиготность
популяции и как следствие – инбридинговая депрессия популяции патогена.
Инбридинг определяется как процент
вероятности для двух аллелей быть идентичными в потомстве. Фенотипический
коэффициент инбридинга (СIph)
может быть рассчитан, по снижению репродуктивной активности патогена или
по снижению уровня развития болезни (формула 3).
СIph = 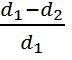 ,
( 3 )
,
( 3 )
где СIph – фенотипический коэффициент
инбридинга, d1 – развитие болезни в открытых насаждениях, d2 – развитие болезни в изолированных на ≈
0,25 га участках насаждений.
Согласно данным таблицы 2, фенотипический
коэффициент инбридинга равен 0,79, что свидетельствует о достаточно высоком
уровне влиянии инбридинга на развитии мучнистой росы.
Прикладной аспект проблемы использования
инбридинга в защите леса от патогенных организмов заключается в том, чтобы
формировать гетерогенные насаждения (Рис. 1, 2). По существу, на основе
предложенных моделей формируется мозаичность насаждений с упорядоченным
размещением парцелл – изолированных деревьев и прогалин.
Парцеллярность определяет мозаичность
лесных экосистем. Мозаичная структура насаждений повышает биоразнообразие и
способствует микоризообразованию. Микоризные древесные более устойчивы к
болезням и неблагоприятным погодным явлениям. Берёза повислая (Betula pendula) способствует распространению
эктомикоризных грибов с широким спектром хозяинных древесных пород. Берёза
способствует улучшению почвы и, формируя ажурную крону, проникновению
рассеянного света в насаждение.
В гетерогенных
дубово-сосново-берёзовых насаждениях более активны процессы естественного
возобновления древесных пород, такие насаждения более привлекательны для
насекомоядных птиц.
Эффективность создания групповых
лесных культур в качестве превентивной лесозащиты подтверждена многолетним
опытом. Так, А.К. Артюховский и др. писали о дубово-сосново-берёзовых
насаждениях, устойчивых к корневой губке (Heterobasidion annosum) [], Б.М. Алимбек рекомендовал
выращивать дуб и интродуцированные древесные породы группами для оздоровления и
повышения продуктивности насаждений. Рекомендуемый нами фенотипический
коэффициент инбридинга позволяет количественно оценивать уровень инбридинговой
депрессии различных вариантов формирования гетерогенных насаждений.
Эффект группы проявляется и в том,
что компактная группа деревьев функционирует как единый, внутренне
взаимосвязанный организм, в котором реализуется естественный отбор.
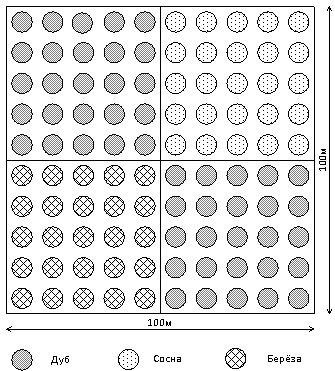
Рисунок 1 – Схема дубово-сосново-берёзовой квадрогруппы
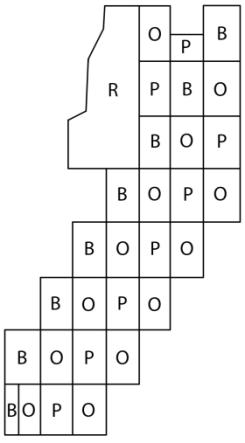
Рисунок 2 – Схема
дубово-сосново-берёзового насаждения
P – Pinus sylvestris (сосна
лесная, O – Quercus robur (дуб черешчатый), В – Betula pendula (берёза
повислая), R – Robinia pseudoacacia (акация)
IV Выводы
Научная новизна изложенного материала заключается в том, что в
результате искусственно вызванного инбридинга в лесных экосистемах реализуется
основополагающий принцип – лучше предотвратить болезнь, чем её лечить.
Инбридинг подавляет популяции патогенных организмов.
Теоретическое значение
исследований определяется новыми данными о сравнительном развитии
мучнистой росы в условиях открытых насаждениях дуба черешчатого и в
экологически изолированных группах деревьев.
Перспективы дальнейших научных разработок в данном направлении
определяются расширением ареала и переходом от формирования гомогенных
насаждений к гетерогенным на эколого-генетической основе.
Литература
1.
Алимбек,
Б.М. Механизированные равномерно-групповые рубки как средство улучшения
состояния и продуктивности дубрав Казанского Поволжья // Состояние и пути
улучшения дубрав РСФСР. Воронеж: ВГУ, 1975. С. 70 – 84.
2.
Артюховский,
А.К. К вопросу создания в очагах корневой губки сосновых насаждений, устойчивых
к грибной инфекции [Текст] / А.К. Артюховский, В.Н. Скрыпников, Ю.Ф. Арефьев //
Сосновые леса России в системе многоцелевого лесопользования: Сб. статей.
Воронеж: ВЛТИ, 1993. С. 76 – 78.
3.
Arefjew Ju.F. Genetisch-ökologische Aspekte des
Forstschutyes. Der Wald, 7’95, Berlin. 1995. S. 238 – 239.