Биология
К. с.-х. н. Седых Т.А., д.б.н. Долматова
И.Ю., д. с.-х. н. Гизатуллин Р.С.
Башкирский государственный аграрный
университет, Россия
Сравнительное
исследование аллелофонда
крупного рогатого скота
герефордской породы
с использованием
микросателитов
Поддержание и
сохранение биологического разнообразия видов сельскохозяйственных животных
является одной из актуальных задач современной науки, решение которой
направлено на повышение жизнеспособности и продуктивности существующих видов, пород,
линий и кроссов, создание новых и п6ерспективных и, в конечном счете, на
успешную реализацию программ по обеспечению населения страны высококачественными
продуктами питания [4,7,9].
В целях реализации программы
развития мясного скотоводства в Республику Башкортостан в 2009 г. был завезен из Австралии крупный рогаты скот герефордской
породы, представленный популяциями, разводимыми в штатах Южная Австралия,
Виктория, Новый южный Уэльс и на о. Тасмания [2.3]. Изучение генетического
потенциала этих животных и сравнительная оценка аллелофонда родителей
австралийской генерации с потомством первого и второго поколения российской
генерации представляет значительный интерес для осуществления селекционно-племенной
работы.
В настоящее время
для объективной характеристики видов и пород сельскохозяйственных животных
широко используются молекулярно-генетические маркеры и одним из
высокоинформативных типов ДНК-маркеров, характеризующих генофонд, являются микросателиты
[5,8].
Целью работы
стало исследование аллелофонда крупного рогатого скота герефордской породы с
использованием микросателитов.
Материалы
и методы исследований. Исследования
проводились на базе ЦКП «Биоресурсы и биоинженерия сельскохозяйственных
животных» ГНУ ВИЖ Россельхозакадемии. Материалом для исследований служили пробы
ткани (ушной выщип) крупного рогатого скота герефордской породы трёх генетико-экологических
генераций в количестве 66 голов, разводимых в МТС «Центральная» Республики
Башкортостан, в т.ч. завезённые из Австралии, (ГРФ_АВСТР, n=30), первой российской генерации,
(ГРФ_F1, n=26) и второй российской генерации (ГРФ_F2, n=10).
Выделение ДНК проводили с помощью набора
реагентов для выделения ДНК DIAtomTM DNA Prep100 (ООО «Лаборатория Изоген»,
Россия). Анализ ДНК и постановку ПЦР проводили по методикам, разработанным
ранее [6]. Набор маркеров для анализа включал 9 микросателлитных локусов -
TGLA126, TGLA122, TGLA227, ILST005, ETH185, ETH10, ETH225, BM1818 и BM2113.
Фрагменты амплификации идентифицировали на генетическом анализаторе ABI Prism
3130xl («Applied Biosystems», США). Статистическую обработку данных проводили
по стандартным методикам [1] с использованием программного обеспечения GenAlEx
v 6.4 [11] и PAST в соответствии с методическими подходами, разработанными в
ГНУ ВИЖ РАСХН [10].
Результаты собственных исследований. Микросателлиты
являются удобными генетическими маркерами в геноме сельскохозяйственных
животных вследствие высокого уровня полиморфизма и стабильного аутосомного кододоминантного
наследования.
|
|
|
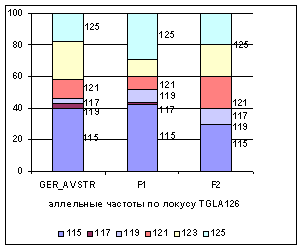
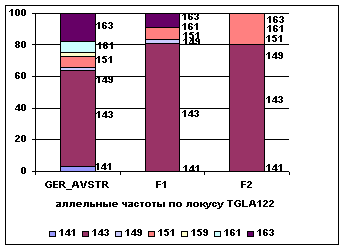
Рис. 1 Аллельные частоты по локусам
TGLA126, TGLA122
Аллельные частоты по микросателлитным
маркерам TGLA126, TGLA122 в разрезе генераций герефордского скота приводятся на
рисунке 1.
Анализ рисунка 1
позволяет утверждать, что по локусу TGLA122, частота аллелей 143 и 151
возрастает от 0,61 и 0,07 у завезённых герефордов (ГРФ_АВСТР) до 0,81 и 0,08 у ГРФ_F1 и 0,80 и 0,2 у ГРФ_РФ_F2, соответственно. По другим аллелям наблюдается снижение частот
и их полное отсутствие во втором поколении. В локусе TGLA126
наблюдается возрастание частоты аллелей 115 и 119 с 0,4 и 0,03 до 0,42 и 0,08 у ГРФ_F1 и 0,3 и 0,1 у ГРФ_РФ_F2, соответственно. Частоты аллелей 121 и 125 в локусе TGLA126
увеличиваются во втором поколении с 0,12 и 0,18 до 0,2, у потомства наблюдается уменьшение частоты аллеля 117, которая
не определена во втором поколении.
На рисунке 2
приводятся аллельные частоты по микросателлитным маркерам TGLA227,
ILST005 в разрезе генераций герефордского скота.
|
|
|
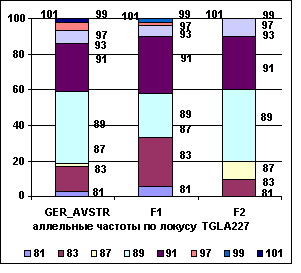

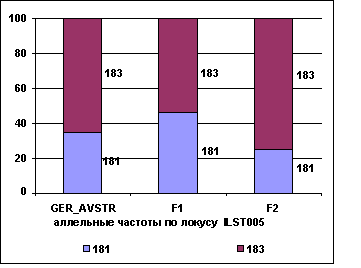

Рис. 2 Аллельные частоты по локусам
TGLA227, ILST005
Полученные данные свидетельствуют о том,
что в локусе TGLA227 частоты аллелей 81 и 83
возрастают от 0,03 и 0,14 у завезённых герефордов (ГРФ_АВСТР) до 0,06 и 0,27 у ГРФ_F1 и снижаются до 0 и 0,10 у ГРФ_РФ_F2, соответственно. У потомков возрастает
частота аллеля 91 - с 0,27
до 0,32 у ГРФ_F1 и до 0,3 ГРФ_РФ_F2. У потомков второго поколения
в локусе TGLA227 отсутствуют аллели 81, 97, 99 и
101. В локусе ILST005 частота аллеля 181 у ГРФ_АВСТР составляет 0,35, увеличивается у потомков ГРФ_F1 до 0,46 и снижается до 0,25 у ГРФ_РФ_F2. Обратная тенденция
наблюдается в этом локусе по частоте аллеля 183 (у ГРФ_АВСТР – 0,65), которая во втором
поколении снижается до 0,54 и в третьем увеличивается до 0,75.
На рисунке 3
приводятся аллельные частоты по микросателлитным
маркерам TGLA227, ILST005 в разрезе генераций герефордского
скота.
|
|
|
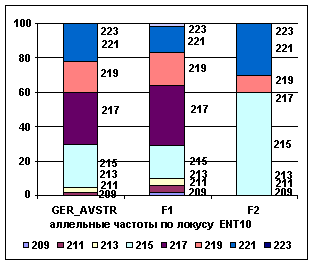

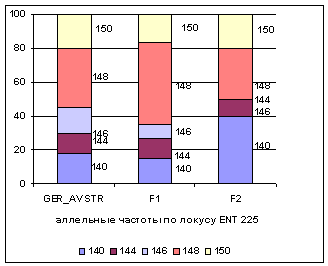
Рис. 3 Аллельные
частоты по локусам ENT10, ENT225
Анализ рисунков позволяет сказать, что в
локусе ENT10 частоты аллелей 215 и 221 у ГРФ_АВСТР находятся на уровне 0,25
и 0,22, снижаются у ГРФ_F1 до 0,19 и 0,15
и возрастают у ГРФ_РФ_F2 до
0,60 и 0,30 соответственно. В локусе ENT10 ГРФ_РФ_F2
не определены аллели 209, 210, 211, 213, 217, 223. В локусе ENT225 наблюдается возрастание частоты аллеля 148 с 0,35 у ГРФ_АВСТР до 0,48 у ГРФ_F1, снижение у потомков первого поколения частот
аллелей 140 и 150 с 0,18 и 0,2 до 0,15 и 0,17, а затем обратное увеличение
показателей до 0,4 и 0,2 у ГРФ_РФ_F2,
соответственно. У потомков второго поколения в локусе ENT225 не определен аллель 146.
На рисунке 4
приводятся аллельные частоты по
микросателлитным маркерам ENT185 в разрезе
генераций герефордского скота.
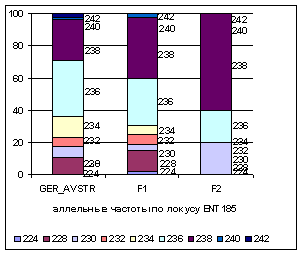
Рис.4 Аллельные частоты в локусе ENT185
В данном локусе у потомков наблюдается
снижение частоты аллеля 230 с 0,07 у ГРФ_АВСТР до 0,04 у ГРФ_F1 и увеличение
до 0,2 у ГРФ_РФ_F2; стойкое
снижение частоты аллеля 236 с 0,35 у ГРФ_АВСТР до 0,29 у ГРФ_F1 и 0,2 у ГРФ_РФ_F2 и увеличение частоты аллеля 238 от 0,25 у ГРФ_АВСТР до 0,38 у ГРФ_F1 и 0,6 у ГРФ_РФ_F2. Во втором поколении в локусе ENT185
не определены аллели 224, 228, 232, 234, 240, 242.
Аллельные частоты по микросателлитным маркерам BM2113 и BM1818 в разрезе
генераций герефордского скота приводятся на
рисунке 5.
|
|
|
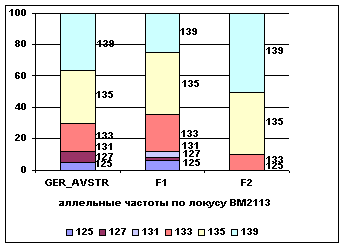
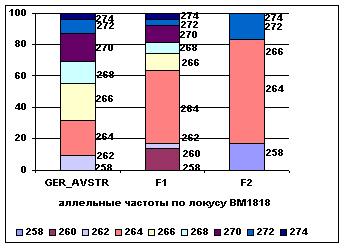
Рис. 5 Аллельные
частоты по локусам BM2113 и BM1818
В локусе BM2113 у потомков наблюдается повышение частот аллелей 135 и 139 с 0,33 и 0,37 у ГРФ_АВСТР до 0,4 и 0,25 у ГРФ_F1 и 0,4 и 0,5 у ГРФ_РФ_F2 и изменение частоты аллеля 133 от 0,18 у ГРФ_АВСТР до 0,23 у ГРФ_F1 и 0,1 у ГРФ_РФ_F2. у потомков второго поколения в данном локусе не определены
аллели 125, 127 и 131. В локусе BM1818 у потомков происходит повышение частоты аллеля
264 от 0,23 у ГРФ_АВСТР до
0,46 у ГРФ_F1 и 0,66 у ГРФ_РФ_F2. У потомков первого поколения
определяется частота аллеля 260 – 0,14, у потомков второго поколения – 258 –
0,17, в локусе BM1818 не определяются аллели 260, 262,
266, 268, 270, 274.
Число аллелей в исследованных локусах варьирует от 2
в локусе ILST005 до 9 в локусах
ETH185, TGLA227 и BM1818. Суммарно в изученной популяции животных выявлено 66
аллелей.
Анализ приватных аллелей, которые
встречаются только в одной из исследованных генераций, показал их наличие во
всех исследованных генерациях животных (рисунок 6).
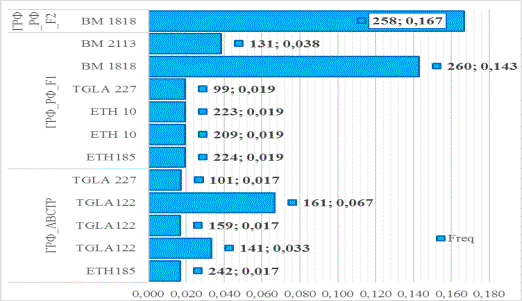
Рис. 6 Частота
встречаемости приватных аллелей в исследованных локусах микросателлитов
различных эколого-генетических генерациях герефордов
Так, в исходной, завезённой генерации
ГРФ_АВСТР в локусе TGLA122, хотя и с низкой частотой, выявлено наличие 3
приватных аллелей (141, 159 и 161). В генерации ГРФ_РФ_F1 приватные
аллели выявлены в 5 локусах, а в генерации ГРФ_РФ_F2 – только в одном- BM1818.
Вывод. На основании проведенных исследований полиморфизма 9 микросателитных
локусов популяции крупного рогатого скота герефордской породы австралийской
генерации и их потомков первого (F1)
и второго (F2) поколений показано,
что в трёх последовательных генерациях животных происходит снижение числа
аллелей во втором поколении. Для более полной характеристики популяций животных
необходимо определить наличие информативных и эффективных аллелей, а также
индекс Шеннона.
С чувством глубокой признательности благодарим
Гладырь Е.А., канд. биол. наук, заведующую лабораторией молекулярной генетики
животных и сотрудников ЦКП «Биоресурсы и биоинженерия сельскохозяйственных животных»
ГНУ ВНИИЖ Россельхозакадемии за помощь, оказанную в проведении исследований.
Литература:
1.
Вейр Б. Анализ
генетических данных // М.: Мир, 1995. 319 с.
2.
Гизатуллин Р.С.
Ресурсосберегающая технология разведения мясного скота и производства говядины:
рекомендации / Р.С. Гизатуллин, Ф.С. Хазиахметов,
Т.А. Седых, Р.М. Мударисов. – Уфа:
Издательство Башкирский ГАУ, 2013. – 64
с.
3.
Гизатуллин Р.С.
Резервы увеличения производства говядины в Башкортостане / Р.С. Гизатуллин, Т,А. Седых
// Вестник
Башкирского государственного аграрного университета. - 2011. - № 3. - С. 25-29.
4. Долматова И.Ю. Зиновьева
Н.А., Горелов П.В., Ильясоа А.Г., Гладырь Е.А., Траспов А.А., Сельцов В.И.
Особенности аллелофонда башкирской популяции симментальского скота по
микросателлитам// Сельскохозяйственная биология. 2011. №6. С. 70-74.
5.
Зиновьева Н.А., Гладырь Е.А. Генетическая
экспертиза сельскохозяйственных животных: применение тест-систем на основе
микросателлитов // Достижения науки и техники АПК. 2011. № 9. С. 46-48.
6.
Зиновьева Н.А.,
Попов А.Н., Эрнст Л.К., Марзанов Н.С., Бочкарев В.В., Стрекозов Н.И., Брем Г.
Методические рекомендации по использованию метода полимеразной цепной реакции в
животноводстве // Дубровицы. ВИЖ, 1998. 47 с.
7.
Зиновьева Н.А., Стрекозов Н.И.,
Молофеева Л.А. Оценка роли
ДНК-микросателлитов в генетической характеристике популяции черно-пестрого
скота // Зоотехния. 2009. № 1. С.
2-4.
8.
Стрекозов
Н.И., Зиновьева Н.А., Горелов П.В., Листратенкова В.И., Коновалова Е.Н.,
Чернушенко В.К., Эрнст Л.К. Генетическая характеристика созданных типов скота
бурой швицкой и сычевской пород с использованием полиморфизма микросателлитных
локусов// Сельскохозяйственная биология. 2009. № 2. С. 10-15.
9. Траспов А.А., Зиновьева
Н.А., Долматова И.Ю., Гладырь Е.А. Аллелофонд башкирской популяции
черно-пестрого скота по микросателлитам в связи с показателями молочной
продуктивности коров // Проблемы биологии продуктивных животных. 2011. Т. 1. С.
65-68.
10.
Nei, M. 1978. Estimation of
average heterozygosity and genetic distance from a small number of individuals.
Genetics 76: 379-390.
11.
Peakall, R., Smouse P.E. GENALEX
6: Genetic analysis in Excel. Population genetic software for teaching and
research. //. Molecular Ecology Notes, 2006, 6, 288-295. V. GENALEX 6.4 UTR: http://appliedpopulationgenetics2010.wikispaces.com/file/view/GENALEX6.4_documntation.pdf,
(дата
обращения
19.12.2010).