Медицина/8. Морфология
УДК 591.446:615.37
К. мед. н. Петизина О.
Н.
ГЗ «Луганский
государственный медицинский университет», кафедра гистологии, цитологии и
эмбриологии, г. Луганск.
ОСОБЕННОСТИ МОРФОЛОГИЧЕСКОГО СТРОЕНИЯ ПОДМЫШЕЧНЫХ ЛИМФАТИЧЕСКИХ УЗЛОВ
КРЫСЫ
Донбасс – это
крупный промышленный регион Украины, в котором насчитывается несколько тысяч
крупных промышленных предприятий, производственно-промышленных объединений и
предприятий топливно-энергетического комплекса, горнодобывающей,
металлургической, химической промышленности, тяжёлого машиностроения,
строительной отрасли, а также агропромышленного комплекса. Донбасс обеспечивает
большую часть промышленного производства Украины, причём в наиболее
экологически опасных отраслях. Высокая концентрация промышленного и сельскохозяйственного производства,
транспортной инфраструктуры, в сочетании с высокой плотностью населения,
создали чрезвычайно высокую техногенную и антропогенную нагрузку на биосферу –
наивысшую в Украине и Европе. Среди экологических факторов,
влияющих на организм человека можно выделить физические, химические и
биологические. Реакции адаптации (приспособления) проявляются на уровне
различных, и в первую очередь, регуляторных систем (нервной, эндокринной,
иммунной). Антропогенные факторы вносят дополнительный вклад и нередко приводят
к срыву адаптационных возможностей организма [10, 20]. С этой точки зрения, иммунная
система является индикаторной системой экологического неблагополучия, поскольку
она чутко реагирует на изменение условий окружающей среды. Одно
из главных мест в иммунной системе занимают лимфатические узлы, в которых
сосредоточены все основные структурно-функциональные компоненты, необходимые
для полноценного иммунного ответа на воздействие внешних факторов [11, 12, 24]. Лимфатические узлы расположены
на пути следования лимфы по лимфатическим сосудам от органов и тканей к
лимфатическим протокам и лимфатическим узлам, таким образом, выполняют функцию биологических фильтров. В них задерживаются любые крупные чужеродные структуры, погибшие
клетки, пылевые частицы и микробные тела, если они оказались в притекающей к
узлу лимфе, а также опухолевые клетки. В синусах лимфатических узлов все эти
структуры задерживаются в петлях сети, образованной ретикулярными волокнами и
клетками, распознаются лимфоцитами и уничтожаются, если их могут поглотить и
«переварить» макрофаги. Такие структуры, как пылевые частицы, захватываются
макрофагами и переносятся в паренхиму лимфатических узлов. Таким образом,
пройдя через лимфатические узлы, лимфа очищается от посторонних частиц и
вливается в кровь уже без чужеродных веществ [15].
Подмышечные
лимфатические узлы крыс относятся к лимфатическим узлам грудной конечности и
туловища (рис. 1.2). Обычно в количестве четырех, лимфоузлы располагаются ниже большой
грудной мышцы на поверхности большой кожной мышцы вдоль стенки грудной клетки, около угла, образованного
грудной стенкой и конечностями. Они собирают лимфу из области грудной
конечности, туловища и принимают лимфу от паховых узлов [13].
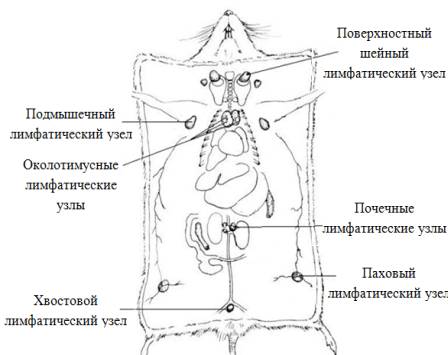
Рис. 1.2.
Схема расположения лимфатических узлов крысы (А. Д. Ноздрачев, 2001).
Лимфатические
узлы крыс представляют собой округлые, мягкие образования, окруженные капсулой.
На срезах узла, проведенных через его ворота, можно различить периферическое,
более плотное корковое вещество, состоящее из лимфатических узелков,
паракортикальную (диффузную) зону, а также, центральное светлое мозговое
вещество, образованное мозговыми тяжами и синусами. Большая часть коркового
вещества и мозговые тяжи составляют область заселения В-лимфоцитов (В-зона), а
паракортикальная, тимусзависимая зона, содержит преимущественно Т-лимфоциты (Т-зона) [13, 15].
Процесс развития
лимфатических узлов у крыс начинается примерно на 11-е сутки эмбриогенеза и
заканчивается через 2 недели после рождения животного.
В развитии лимфатических
узлов выделяют несколько стадий:
1)
образование
лимфатических мешочков [22];
2)
деление
и почкование клеток эндотелия – образование лимфатических сосудов;
3)
формирование
инициирующего зачатка лимфатического узла;
4)
образование
субкапсулярного синуса и заселение лимфатического узла клетками гемопоэтических
предшественников;
5)
образование
капсулы органа и миграция зрелых лимфоцитов.
В развитии зачатков
лимфатических узлов принимают участие две популяции клеток: индуцирующие клетки
(LTIC – lymphoid tissue inducing cells) и клетки-организаторы. Взаимодействие
этих двух видов клеток приводит к индукции экспрессии соответствующих лигандов
и рецепторов на их поверхности, а также к секреции хемокинов, способствующих
направленной миграции дополнительных клеток в развивающийся орган [21, 22].
Таким образом, для
начала развития изучаемого органа необходимо оптимальное соотношение
стимулирующих сигналов со стороны как индуцирующих, так и стромальных клеток, и
даже незначительное нарушение этого баланса приводит к нарушению развития
лимфатических узлов, повлиять на которое во взрослом состоянии уже невозможно [7,
19].
Кроветворная функция
лимфатических узлов заключается в антигензависимой пролиферации и
дифференцировке Т- и В-лимфоцитов. Кроме того, выполняя защитную функцию,
лимфатические узлы участвуют в иммунных реакциях. Проходя сквозь них, лимфа
очищается от чужеродных частиц, микроорганизмов, а также обогащается
лимфоцитами и иммуноглобулинами [17].
Основной каркас
подмышечного лимфатического узла крыс образован соединительнотканной капсулой и
отходящими от неё в
паренхиму узла трабекулами. Паренхима органа, состоящая преимущественно, из
лимфоцитов, подразделяется на корковое, мозговое вещество и паракортикальную
зону. Клетки стромы в разных участках узла отличаются по морфологическим
критериям. В областях,
богатых В-лимфоцитами, например, в лимфатических узелках, преобладают
дендритные клетки, их цитоплазма разветвляется и образует много выростов. Они
не фагоцитируют, но могут связывать на своей поверхности
антигены и вовлекать в иммунную реакцию контактирующие с ними В-лимфоциты [2,
3, 5, 23]. В зонах богатых Т-лимфоцитами, например, в паракортикальной зоне,
клетки стромы имеют полиморфное ядро и сплетающиеся длинные пальцевидные
отростки. Они носят название «интердигитирующие клетки». Данные клетки являются
своеобразной системой, способной воспринимать и передавать антигенную
информацию на иммунокомпетентные клетки. Также строма богата клетками с
признаками макрофагов. Между соединительнотканным каркасом и паренхимой имеются
выстланные литоральными клетками пространства – синусы: краевой,
вокругузелковый, мозговой и воротный [8, 15, 16].
На гистологических
препаратах лимфатических узлов крыс корковое вещество занимает большую площадь
среза. Оно имеет неоднородное строение. На микрофотографиях коркового вещества
отчетливо видны округлой формы – лимфатические узелки, размером 0,5 – 1 мм в
диаметре. Большинство авторов выделяют первичные и вторичные лимфатические
узелки. В строении узелков различают герминативный центр, в котором располагается множество делящихся и молодых клеток
лимфоидного ряда, в основном В-лимфоцитов, являющихся предшественниками
антителообразующих клеток – плазмоцитов, типичных макрофагов, дендритных
ретикулярных клеток и лимфоцитов с повышенной активностью. Периферическая часть
лимфоидных узелков называется короной и содержит большое количество малых и
средних В-лимфоцитов [1, 9, 23]. Паракортикальная зона располагается между
корковым и мозговым веществом и состоит из диффузно расположенных Т-лимфоцитов
и интердигитирующих клеток. Мозговое вещество образовано преимущественно
В-лимфоцитами и плазмоцитами, расположенными в мозговых тяжах, которые
представляют собой петли ретикулярной ткани, образованные ретикулярными
клетками и волокнами [4, 19].
Первичный лимфатический
узелок лимфатических узлов крыс образован преимущественно лимфоцитами. Их ядра
крупные, расположены в центре. В нуклеоплазме, в основном, выявляется
эухроматин, гетерохроматин занимает узкую полоску под внутренней ядерной
мембраной. Иногда выявляется ядрышко, имеющее зернистую структуру. Среди
лимфоцитов этого типа встречаются клетки, кариолемма которых имеет одно или
несколько вдавлений, в кариоплазме встречаются более массивные скопления
гетерохроматина. Цитоплазма клеток узкая, часто плазмолемма формирует
микроворсинки и углубления, которые придают лимфоцитам неправильную форму. В
цитоплазме идентифицируются многочисленные свободные рибосомы и полисомы,
овальные митохондрии, гранулярная эндоплазматическая сеть, умеренно развитый
комплекс Гольджи [1, 19, 22]. Дендритные ретикулярные клетки первичных лимфатических узелков
имеют хорошо развитые цитоплазматические отростки и электронноплотную
цитоплазму. Эти клетки образуют по периферии узелка мелкопетлистую сеть.
Вторичный лимфатический
узелок коркового вещества лимфатических узлов крыс образован различными типами
лимфоцитов (малыми, средними, большими, лимфобластами). Очень редко в составе
вторичных лимфатических узелков обнаруживаются макрофаги, строма представлена
дендритными клетками. Благодаря наличию отростков они контактируют между собой
и многочисленными лимфоцитами. Тесные межклеточные контакты стромальных клеток
с лимфоцитами узелков являются структурно-функциональной основой иммунного
ответа по гуморальному типу в В-зависимых зонах лимфатических узлов. Даже в
пределах одного лимфатического узелка дендритные клетки отличаются по
морфологической характеристике, способу распределения хроматина в ядре, количеству,
степени развитости органелл, плотности и распределению включений в цитоплазме [5,
17]. Типичным признаком субмикроскопической организации дендритных клеток
является отсутствие крупных вторичных лизосом, гетерофагосом, поскольку они
обладают очень низкой фагоцитирующей
активностью [24].
Среди компонентов стромы
соматических лимфатических узлов присутствуют фибробластические ретикулярные клетки.
Они характеризуются вытянутой формой, имеют отростки, в
цитоплазме
содержат крупное овальное ядро с ядрышком. В нуклеоплазме преобладает
мелкодиспергированный эухроматин. Особенностью цитоплазмы этих клеток является
наличие сильно развитых канальцев гранулярной эндоплазматической сети и
комплекса Гольджи, что свидетельствует о высокой синтетической способности фибробластических
ретикулярных клеток, играющих ведущую роль в выработке межклеточного вещества и
ретикулярных волокон [23].
Паракортикальная зона
содержит многочисленные Т-лимфоциты с типичной ультраструктурой. Они
расположены в окружении клеток ретикулярной стромы [3, 18]. Для данной зоны
лимфатических узлов интактных крыс характерно наличие интердигидирующих
ретикулярных клеток, имеющих моноцитарное происхождение. Ядра их неправильной
формы с глубокими инвагинациями кариолеммы, содержат эухроматин. Цитоплазма
характеризуется низкой электронной плотностью с развитой эндоплазматической
сетью, представленной гранулярными и гладкими узкими профилями. Многочисленные
митохондрии, разные по величине, округлой формы с матриксом умеренной
электронной плотности, четко контурированными и упорядоченно расположенными
кристами, локализуются преимущественно около ядра. Интердигитирующие клетки
имеют длинные пальцевидные цитоплазматические отростки. Для паракортикальной
зоны лимфатических узлов крыс характерным является наличие в ней большого
количества посткапиллярных венул с высоким эндотелием, через которые
осуществляется миграция лимфоцитов, как в паренхиму узла, так и в
противоположном направлении – в кровь [19, 22].
Мозговое вещество лимфатических
узлов в целом является В-зависимой структурнофункциональной зоной. При анализе
электронограмм мозгового вещества подмышечных лимфатических узлов крыс
идентифицируются лимфоциты типичной ультраструктуры, входящие в состав мозговых
тяжей, здесь находятся также плазматические клетки и
макрофаги [6,14].
Сделав анализ
вышеизложенного материала, можно сделать вывод, что изучение подмышечных
лимфатических узлов является актуальным и представляет большой интерес для
современной науки.
Литература:
1. Абрамова М. В. Динамика
клеточного состава брыжеечных лимфатических узлов мыши после их облучения
изотопом 131Сs / М. В. Абрамова, П. Г. Магомедова // Морфология. – 2010. –
Т. 137, №
4. – С. 12–13.
2. Абрамова М. В.
Морфологические аспекты изменения структурной организации и клеточного состава
брыжеечных лимфоузлов белых мышей, облученных потоком гамма-частиц / М. В.
Абрамова, П. Г. Магомедова // Морфология. – 2008. – Т. 133, № 2. –
С. 6.
3. Александрова Л. И.
Планиметрическая характеристика лимфатических узлов при экспериментальном
воздействии переменного электромагнитного поля промышленной частоты / Л. И.
Александрова, М.
Ю. Капитанова, Н. Г. Краюшкина // Морфология. – 2010. – Т. 137, № 4. – С. 14.
4. Ноздрачев
А. Д. Анатомия крысы (Лабораторные крысы) / Ноздрачев А. Д., Поляков
Е. Л. – Санкт-Петербург: Издательство «Лань», 2001. – 464 с.
5. Ахадова З. А.
Гистологические показатели шейных лимфатических узлов / З. А. Ахадова //
Морфология. – 2010. – Т. 137, № 4. – С. 21.
6. Бахмет А. А. Морфологические особенности некоторых органов иммунной системы
крыс Вистар при стрессе и воздействии пептида, вызывающего дельта-сон / А. А.
Бахмет, Е. В. Коплик // Морфология. – 2007. – Т.
131, № 3. –
С. 58.
7. Бахмет А. А. Морфологическая характеристика селезенки, паховых
лимфатических узлов и лимфоидных бляшек тонкой кишки крыс при эмоциональном
стрессе, а также в
условиях воздействия некоторых олигопептидов: автореф. дис. на соиск. ученой
степени доктора мед. наук, спец. 14.03.01 «нормальная анатомия» / А. А. Бахмет. – Москва, 2010. –
46 с.
8. Волкова Л. В.
Акцидентальная инволюция лимфоидных органов и оценка межклеточных
взаимодействий / Л. В Волкова //
Морфология. –
2007. – Т. 131, № 3. – С. 62.
9. Головацкий А. С. Особенности
рециркуляции лимфоцитов через посткапилярные венулы паракортикальной зоны
лимфатических узлов при антигенной стимуляции / А. С. Головацкий, М. Р. Сапин
// Морфология. – 2001. – Т.
119, № 1. – С. 40–45.
10. Григоренко Д. Е.
Морфофункциональное состояние печеночных лимфатических узлов при
экспериментальном гепатите / Д. Е. Григоренко, М. Р. Сапин, А. М. Хребтовский
// Вестник новых медицинских технологий. – 2006. – Т 13, № 1. – С. 30–33.
11. Гусейнов Т. С. Влияние
гидрологических факторов на морфогенез иммунных органов / Т. С. Гусейнов, С. Т.
Гусейнова // Морфология. – 2007. – Т. 131, № 3. – С. 65.
12. Давтян Т. К. Эволюция
интегративной функции иммунной системы / Т. К. Давтян, Г. А. Геворкян, Д. А.
Погосян // Успехи современной биологии. – 2005. – № 1. – С. 34–40.
13. Дмитриев Д. А.Современные методы
изучения влияния загрязнения окружающей среды на иммунную систему / Д. А. Дмитриев, Е. Г. Румянцева // Гигиена и
санитария. – 2002. – № 3. – С. 68–71.
14. Ноздрачев
А. Д. Анатомия крысы (Лабораторные крысы) / Ноздрачев А. Д., Поляков
Е. Л. – Санкт-Петербург: Издательство «Лань», 2001. – 464 с.
15. Пинегин Б. В.
Нейтрофилы: структура и функция / Б. В. Пинегин, А. Н. Маянский // Иммунология.
– 2007. – № 6. – С. 374–382.
16. Сапин М. Р. Лимфатический узел /
Сапин М. Р., Юрина Н. А., Этинген Л. Е. – Москва: Медицина, 1978. – 272
с.
17. Сепиашвили Р. И. Апоптоз в иммунологических процесах / Р. И.
Сепиашвили, М. Г. Шубич // Аллергология и иммунология. – 2000. – № 1. – С.
15–23.
18. Субпопуляции
Т – лимфоцитов в центральних и периферических лимфоидных органах при воспалении
/ Т. И. Дергачева, А. В. Шурлыгина, А. В. Юкляева [и др.] // Імунология. –
2009. – Т. 30, № 2. – С. 101–104.
19. Суховершин, А. А. Региональные лимфатические узлы конечности при
воспалении / А. А. Суховершин, О. В. Казаков, В. В. Асташов // Морфология. – 2008. – Т. 133, № 2. – С. 131.
20. Танасийчук И. С. Цитоморфологическая характеристика клеточного состава
лимфатических узлов в норме / И. С. Танасийчук // Цитология и генетика. – 2004.
– № 6. – С. 60–66.
21. Трахтенберг И. М. Экологическая безопасность – глобальная
социально-медицинская проблема / И. М. Трахтенберг // Гигиена и санитария. –
2009. – № 7 (63). – С. 32–37
22. Туманов
А. В. Развитие вторичных лимфоидных органов / А. В. Туманов // Иммунология. –
2004. – № 2. – С. 120–128
23. Anderson
A. O. Conduit for Privileged Communications in the Lymph Node / A. O. Anderson,
S. Shaw // Immunology. – 2005. – Vol. 22, № 1. – Р. 3–5.
24. Balogha P. Fibroblastic
reticular cells of the peripheral lymphoid organs: unique features of a
ubiquitous cell type / P. Balogha, V. Fisia, A. K. Szakalb //
Molecular Immunology. – 2008. – Vol. 46, № 1. – P. 1–7.
25.
Lymphatic system: morphofunctional
considerations / G. Sallustio, C. Giangreogorij, L. Cannas [et al.] // Rays. – 2000. – Vol. 25, № 4. – P 413–427.