ФИЗИЧЕСКАЯ ПРИРОДА МЕХАНИЗМОВ ЯВЛЕНИЯ ИНДУКЦИИ
А.В. Бобров
Орловский Государственный университет –
учебно-научно-производственный комплекс.
Россия
Ключевые слова: индукция, структура, топологические
параметры, информационное воздействие, информационная матрица, взаимодействие
собственных спиновых полей
Показана физическая природа обнаруженного
в 1901 году явления Индукции (Явления информационного
взаимодействия собственных спиновых полей материальных объектов). Механизмы
Индукции основаны на свойстве собственных спиновых полей материальных
объектов непосредственно Взаимодействовать
в пределах ближней зоны. Такие взаимодействия, не нарушая структуру вещества,
влияют на состояние спиновых полей, что обуславливает изменение свойств
вещества, и в итоге приводит к изменению физических, физико-химических и
биологических процессов, в которых объекты участвуют. Механизмы Индукции контролируют развитие живого многоклеточного
организма и его жизнедеятельность на протяжении всей жизни.
Собственные спиновые поля материальных объектов (ССП МО)
несут информацию о структуре вещества и используются в медицине для визуализации
внутренних структур и органов человека методом магнитно-резонансной томографии.
Согласно результатам наших исследований, собственные спиновые поля
"шагнули" за пределы материальных объектов.
В 2010 году было
обнаружено явление Информационного взаимодействия ССП МО, основанное на их специфических
свойствах:
1. Нести информацию о геометрии и
топологии материального объекта, структуре его вещества и его элементном
составе.
2. Взаимодействовать непосредственно
(спин-спиновые взаимодействия) при удалении материальных объектов друг от друга
на расстояние, не превышающее протяженность "ближней зоны", которая
зависит от различных параметров взаимодействующих объектов и может доходить до
2-3 м.
3. взаимодействовать опосредованно при
участии дальнодействующих спиновых полей Физического Вакуума в случае, если
расстояние между взаимодействующими материальными объектами превышает
протяженность "ближней зоны..
4. Воздействовать на физические,
физико-химические и биологические процессы, происходящие в материальных
объектах изменяя при этом их свойства. [1]-[2].
Открытие
явления информационного взаимодействия повлекло за собой определение механизмов и физической природы феномена Индукции [3], “зависшего” в биологической науке не
разрушаемым “белым пятном” более чем на целое столетие.
Первые многоклеточные организмы возникли
более миллиарда лет назад в результате включения Собственных спиновых полей в
процессы эволюционного развития живых систем. Наука впервые познакомилась с их
проявлением в 1901 году, когда немецкий эмбриолог Х. Шпеман, изучая образование
хрусталика глаза у зародышей земноводных, обнаружил феномен Индукции (Нобелевская
премия 1935 г). За всё последующее столетие эмбриологи наработали по феномену
Индукции богатейший экспериментальный материал, но не приблизились к пониманию
его природы и механизмов. Решение этих проблем стало возможным только после
обнаружения в 2010-м году явления Информационного взаимодействия ССП МО.
Результат проявления феномена
Индукции легко доступен для понимания: "всё в организме влияет на
всё" – все клетки и их компоненты завязаны в едином процессе взаимодействия. Роль Индукции особенно важна для организма, начинающего свое
развитие. Индукция контролирует строительство всех тканей и органов, назначая
каждой вновь возникшей клетке t` локализацию и определяя её функцию в процессах превращения
эмбриона в зрелый многоклеточный организм. Можно с полной уверенностью
утверждать: не будь Индукции, мы все до сегодняшнего дня оставались бы
одноклеточными амёбоподобными организмами. Значимость обнаруженного в начале ХХ
столетии явления Индукции можно сравнить со значимостью обнаруженных в середине
этого же столетия двойной спирали ДНК и
генетического кода. Образно говоря, если ДНК – программа построения будущего
организма, то Индукция это – прораб-строитель, реализующий генетическую программу.
В процесс детерминации, проходящий на начальных этапах развития многоклеточных
тканей (бластуляции, гаструляции), вовлечен механизм феномена Индукции.
Свойства феномена Индукции – память и структурообразование, экспериментально
проявляются уже на уровне цитоскелета. Они проявляются и далее в процессах
дифференцировки на стадиях нейруляции и органогенеза при развития возбудимых
тканей в ЦНС и на периферии, например, при навигации кончика роста аксона в
поиске мишени,
Обширный экспериментальный
материал - адаптированные фрагменты из
источников [4]-[6] позволяет определить основные признаки физического фактора,
реализующего проявления феномена Индукции и его природу, как в процессах естественного
онтогенеза, так и в реакциях, возникающих в ответ на внешние воздействия,
например, при трансплантацию индукторов. Основываясь на эти признаки, можно
утверждать, что Явление Индукции не
просто похоже на явление Информационного взаимодействия ССП МО, а представляют
с ним единое целое. Специфика информации,
характеризующей проявления этого феномена, заключается в привлечении, вместо
привычных дискретных количественных значений поля, иных – качественных
категорий – структурных и топологических показателей. Известными физическими
полями, удовлетворяющими указанному условию, являются несущие информацию
Собственные спиновые поля материальных объектов.
1.
Проявление феномена Индукции на уровне цитоскелета
А. Информация о
структуре цитоскелета материнской клетки может передаваться непосредственно
цитоскелету дочерних клеток. Организация и
функционирование цитоскелета практически не зависят от клеточного ядра. Клетка,
утратившая ядро, все еще способна прикрепляться к субстрату, изменять форму,
мигрировать, захватывать пищевые частицы и т. д. Если отделить от фибробласта
небольшие фрагменты, вокруг каждого из них замыкается плазматическая мембрана
и эти фрагменты сохраняют различные формы подвижности в течение нескольких часов.
Некоторые из них образуют мембранные выпячивания или ламеллоподии, другие вытягивают и втягивают микрошипы. Ни один из них не
способен передвигаться, и это позволяет
думать, что субструктуры цитоскелета сами по себе могут обеспечить только локальную подвижность, тогда как
перемещение всей клетки требует
некоторой согласованности различных двигательных процессов. Можно уверено утверждать, что описанные выше
формы подвижности новообразованных структур никак не связаны с генетической
памятью, поскольку цитоскелет не содержат в себе генетического аппарата. Подтверждением того, что цитоскелет и его фрагменты
обладают своеобразной «памятью», являются сходные мембранные выпячивания, а также сходство в траекториях миграции у двух дочерних
клеток, возникших при делении культивируемой
клетки.
Наиболее четкое свидетельство существования цитоскелетной наследственности
получено в
опытах с крупным одноклеточным организмом Pаrате-сшт. Его поверхность покрыта рядами подвижных ресничек, обладающих свойством
синхронизированного однонаправленного биения. В эксперименте этот строгий порядок
можно нарушить и получить инвертированные ряды, биение ресничек в которых будет
направлено в обратную
сторону. Такая измененная организация передаётся
и последующим поколениям данной клетки непосредственно через цитоскелет, лишённый генетического аппарата.
Итак,
цитоскелет материнской клетки может
передавать информацию об особенностях своей структурной организации
непосредственно цитоскелету дочерних клеток. Как именно передается информация от клетки к
клетке не известно. [4]. Т3,
с. 132-133
Б.
Информация о структурной организации цитоскелета может передаваться за пределы клеточной мембраны. Цитоскелет данной клетки может влиять на
цитоскелет не только дочерних клеток, но и соседних клеток той же ткани. Развитие цитоскелета в соседних клетках не происходит независимо: имеет
место взаимное влияние. Цитоскелеты отдельных клеток, составляющих данную ткань,
во многих случаях организованы таким образом, что напоминают скорее составные части
единой системы, чем независимые структурные единицы. [4]. Т3, с. 131.
2.
Собственное спиновое поле клетки является носителем её памяти
При возникновении
двух первых клеток из оплодотворённой материнской клетки каждая из них «знает»
о существовании другой. У лягушки после первого дробления клетки равноценны и
если их разнести, то из каждой разовьется нормальный головастик, который затем
превратится в лягушку. Но если одну из клеток убить и удалить, то из оставшейся
живой клетки начнет развиваться только половина тела зародыша [4]. Объяснить
этот феномен можно нарушением стационарного индукционного процесса
взаимодействующих ССП двух клеток. Разрушение одной из них, или её
удаление, ведёт к нарушению этого процесса, что влечёт за
собой изменение состояния спинов ССП второй клетки и, соответственно, нарушение
процесса нормального развития второго эмбриона
3. Индукционные взаимодействия при развитии эпителиев
3.1. Мезодерма индуцирует образование различных частей
нервной трубки из эктодермы
Влияние одной ткани на дифференцировку клеток другой ткани, тесно
контактирующей
с первой, является классическим примером проявления феномена Индукции. Многие органы состоят
из двух или большего числа различных клеточных популяций, которые возникли независимо
друг от друга, а позже объединились и стали взаимодействовать между собой.
Дифференцировка клеток в одной ткани может определяться влиянием индукционного
воздействия другой ткани, тесно контактирующей с первой. Подобного рода, взаимодействия играют особенно важную
роль в развитии эпителиев у позвоночных
животных.
Эксперименты на амфибиях показали, что образование нервных структур из эктодермы обусловлено индуцирующим воздействием подстилающей
мезодермы. Если на стадии гаструлы из участка, расположенного под будущей
нервной трубкой одного зародыша,
взять кусочек мезодермы и имплантировать его под эктодерму другого зародыша на брюшной стороне, то эктодерма здесь начнет утолщаться и сворачиваться, образуя в этом
необычном месте отрезок нервной
трубки. Специфика растущей ткани этого отрезка будет зависеть от происхождения трансплантированной мезодермы. Если
мезодерма была взята из переднего
участка, из эктодермы образуется часть головного мозга. Мезодерма, взятая из заднего участка, вызовет
образование отрезка спинного мозга.
При этом специфика растущей ткани на этом отрезке будет зависеть от происхождения трансплантированной мезодермы. Этот результат свидетельствует о том, что клетки
эктодермы приобретают информацию о структуре спиновой системы и топологии, зависящей от позиционных значений
подстилающих клеток трансплантированной мезодермы. На более поздних
стадиях развития дифференцировка многих эпителиальных структур в основном также контролируется подстилающей мезенхимой. Именно таким образом из компонентов эпителия и
мезенхимы образуются кожа и кишка со
всеми специализированными элементами и
производными. [5], Т4, с.
111
3.2 Характер и
распределение производных эпидермиса
контролируется дермой
Кожа
состоит из двух слоев - эпидермиса (эпителий, образующийся из эктодермы) и
дермы (соединительная ткань, образуемая главным образом мезодермой). Из
эпидермиса формируются кератинизированные придатки кожи (волосы, перья, чешуя и
когти), а также многие железы. Для разных участков тела характерны различные
виды кератинизированных производных: на спине и крыльях у цыпленка образуются перья,
а на ногах чешуйки. При этом на спине основания перьев располагаются в виде упорядоченной
гексагональной структуры и образуют ясно выраженные
цепочки.
Если у куриного эмбриона взять
эпидермис с ноги, где он позже образует
чешуйки, и объединить
с дермой спины - области, где в норме вырастают перья, то из него вместо чешуек будут формироваться перья. [5], Т4,
с. 111
3.3. Межклеточные взаимодействия контролируют развитие
желез и
их функцию
Большая часть желез формируется путем врастания эпителия
в подстилающую мезенхиму. Каждый тип желез характеризуется определенной геометрической формой и химизмом
секреции. Например, молочная железа возникает в виде эктодермального зачатка, который погружается в мезенхиму и неоднократно
разветвляется, образуя "дерево" протоков; на концах протоков появляются
особые эпителиальные клетки, которые после соответствующей гормональной стимуляции начнут выделять молоко,- единственные клетки тела, способные выполнять эту функцию. Для
развития железы необходимы индуцирующие сигналы от клеток мезенхимы, однако
интерпретация этих сигналов зависит также от природы реагирующего эпителия. На
ранних стадиях образования различных желез можно
разделять будущие эпителиальные и мезенхимные элементы этих желез и снова объединять их в различных комбинациях in vitro. Такой подход позволяет изучать межклеточные взаимодействия,
контролирующие развитие желез. При объединении эпителия молочной железы с
мезенхимой слюнной железы образующиеся протоки ветвятся по типу слюнной железы.
Однако если
такую железу подвергнуть гормональному воздействию путем имплантации беременной самке, то эпителиальные
клетки, выстилающие концы протоков, начинают
вырабатывать молоко, т.е. секрет, соответствующий их происхождению. [5], Т4, с. 112
4.
Навигация конусов роста в спинном мозге
Рассмотрение механизма
навигации к своим клеткам-мишеням аксонов комиссуральных интернейронов спинного мозга,
подтверждает концепцию информационного взаимодействия спиновых полей
материальных объектов. Этот механизм демонстрирует возможность реализации
уникального свойства взаимодействия спиновых полей – обеспечение процесса
перемещения и сборки компонентов живого организма в единое целое.
В раннем периоде
развития комиссуральные интернейроны, которые лежат в дорзальной части спинного мозга,
выпускают аксоны, которые начинают расти в вентральном направлении, пересекая
среднюю линию, и затем растут вдоль спинного мозга по направлению к своим
синаптическим мишеням.
У позвоночных и беспозвоночных ведущий
край аксона - конус роста, перемещается на значительные расстояния с высочайшей
точностью. Конус роста ведет аксон, действуя как совершенный сенсор, который
воспринимает и соответственно отвечает на различные средовые сигналы наведения.
Эти сигналы существуют или в диффузионной форме или в формах, ассоциированных с
поверхностью клеток, и отвечают, соответственно, за нахождение путей на длинное
и короткое расстояние. Рецепторы клеточной поверхности, располагающиеся на
конусах роста, распознают эти сигналы как позитивные - аттрактивные, или
негативные – репульсивные силы. Дальнодействующие репульсивные наводящие
сигналы, обозначаемые как хеморепелленты, заставляют конусы роста отодвигаться
от препятствия, тогда как дальнодействующие аттрактивные сигналы должны притягивать аксоны на большом расстоянии.
С другой стороны, контактзависимые
аттрактивные и репульсивные сигналы контролируют направленность роста аксонов
на коротких расстояниях. Совместно, все четыре силы наведения формируют
полностью траекторию пути данного аксона. Сложные траектории аксонов разделены
на серии коротких
сегментов. "Промежуточные мишени" маркируют конец одного
сегмента и начало следующего. Большинство промежуточных мишеней представляет собой
небольшие группы специализированных указывающих клеток, которые предоставляют
конусам роста информацию, необходимую для направления перемещения аксона вдоль
следующего сегмента его траектории. В соответствии с их ролью, наблюдаются
резкие изменения в морфологии конусов роста и направления роста аксона, когда
аксон перемещается в непосредственной близости
к промежуточным мишеням, или в некоторых случаях, после непосредственного
контакта с ними. При удалении
промежуточных мишеней, нахождение аксонами пути нарушается.
Срединная линия развивающейся нервной
системы является важной точкой выбора направления движения аксонов. Большинство
аксонов проходит значительные расстояния в направлении срединной линии.
Достигнув её, они должны решить, пересекать её или нет, а также решить, расти
далее в ростральном или каудальном направлении параллельно срединной линии. Оба
класса аксонов ограждены от срединной линии, когда начинают расти в продольном
направлении. Навигация аксонов в этой области обеспечивается способностью
конусов роста правильно интегрировать разнообразные внеклеточные сигналы наведения.
Аксоны нейронов первоначально привлекаются к вентральной средней линии белком нетрин-1 – растворимым хемоаттрактантом, образуемым специальными клетками базальной пластинки, лежащими вдоль средней линии спинного мозга. Нетрин-1 взаимодействует с рецептором, экспрессируемым комиссуральными
нейронами, называемым DCC (рис.1 А).
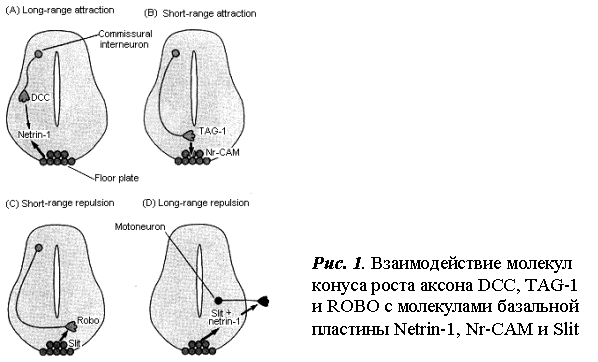
Существование
растворимого
фактора, образующегося в базальной пла-
стинке, который
способен привлекать аксоны комиссуральных нейронов, было
впервые показано in vitro при культивировании кусочков дорзальной
части спинного мозга изолированно, либо вместе с кусочками базальной пластинки. Аксоны комиссуральных
нейронов растут четко по направлению к базальной пластинке, даже если эксплантаты находятся
друг от друга на расстоянии нескольких сотен микрон. Далее аксоны комиссурального нейрона пересекают
вентральную среднюю линию, что они делают только однажды, таким образом, оставаясь на
контралатеральной стороне. Это пересечение облегчается взаимодействиями между двумя молекулами клеточной поверхности: TAG-1, экспрессируеvjq на поверхности аксона, и NrCAM, экспрессируемой на клетках базальной пластинки (рис. 1В).
После пересечения средней линии экспрессия TAG-1
в аксонах ингибируется сигналами от клеток базальной пластинки, под действием которых в
аксонах начинает синтезироваться протеин Кobo. Robo представляет собой рецептор для другого белка, называемого Slit, который
высвобождается клетками базальной пластинки.
Взаимодействие между Slit и Robo отталкивает конусы роста комиссуральных интернейронов
(рис. 1С ). Потеря аттракции к TAG- – NrCAM контакту и приобретение короткодистантного отталкивающего взаимодействия Slit-Robo предупреждают возможное повторное
пересечение аксоном средней линии. Slit и Нетрин-1 также диффундируют из базальной пластинки для
отталкивания конусов роста мотонейронов. Такие отталкивающие взаимодействия направляют аксоны мотонейронов
прочь от спинного мозга в направлении к
периферии (рис. 1D). [6], с. 550-553
Кроме
рассмотренного выше индукционного механизма навигации конуса роста аксона, в
котором взаимодействие белковых дальнодействующих молекул аттрактантов и
реппелентов происходит на большом расстоянии, существует предложенный
Рамон-Кахалем на основании экспериментов in vitro механизм
хемоаттрактантного управления
ростом аксона, согласно которому конус
роста аксона следует по градиенту
концентрации молекул, секретируемых в раствор клетками мишени. Считается, что взаимодействия, основанные
на концентрационном градиенте, возможны лишь при расстоянии нескольких сотен
мкм. Однако концепция механизма
хемоаттрактантного управления ростом аксона не состоятельна. Главным аргументом её несостоятельности служит
возможность перемещения конуса роста аксона in vivo в условиях, при которых секретирование
молекул в раствор не происходят и, следовательно, градиент их концентрации
межклеточной жидкости заведомо отсутствует. На рис. 1В показан результат взаимодействия
конуса роста аксона с базальной пластинкой, возникающий в результате аттракции молекул TAG-1молекулами Nr-CAM, входящими в состав клеток базальной пластинки,. Но молекулы Nr-CAM, также, как и встроенные
в мембрану молекулы ТAG-1, относятся к типу дальнодействующих трансмембранных молекул (рис.2), и не секретируются во внеклеточную
среду, и следовательно, не могут образовывать градиенты концентрации раствора
во внеклеточной среде un vivo. Между тем, несмотря на
отсутствие в ней самих молекул, их воздействие приводит к пересечению
конусом роста вентральной средней линии.
5. Структурный фактор
Информационную
природу в явлении навигации конуса роста аксона подтверждает участие в нём
взаимодействующих белковых молекул, принадлежащих различным семействам
молекул-гомологов, выполняющих в зависимости от структуры прямо противоположные
функции притяжения и отталкивания, обусловленные взаимодействием Собственных
спиновых полей этих молекул.
На рис. 2
показана специфика структуры молекул клеточной адгезии Nr-CAM, TAG-1 и DCC, участвующих в процессе навигации конуса роста аксона. Молекулы представляют собой трансмембранные или связанные с мембраной гликопротеины со сложной структурой своих внеклеточных частей, включающих 4-6 повторяющихся доменов
фибронектина и доменов, которые во многом схожи с постоянными доменами иммуноглобулинов и фибронектина типа 3. Для сравнения на рис. 3
приведена структура трансмембранных молекул
хемореппелентов из семейства семафорины, содержащих во внеклеточной области
домены, напоминающие иммуноглобулины.
К концу прошлого столетия определилось приоритетное
направление в ис-
следовании механизма Индукции – изучение
свойств молекул со сложной структурой и их подбор. Было обнаружено, что
структура сложных белковых молекул предопределяет
их функциональные свойства. Понимание роли структуры стало ключом, позволившим
проведение целенаправленного изучения феномена Индукции, поскольку возникла возможность предсказания
результатов планирующегося эксперимента. Наука ХХ столетия оказалась на пороге
раскрытия механизма и природы феномена Индукции. Однако это понимание не
привело к решению проблемы. Интрига
феномена Индукции оставалась заключенной за семью замками в ожидании
обнаружения явления Информационного взаимодействия Собственных спиновых полей
материальных объектов, основанного на неизвестном ранее явлении
распространения этих полей за пределы самих объектов.

Механизмы Индукции участвуют в процессах
восстановления функции передачи нервного импульса при нарушении целостности нервных волокон мотонейронов и нарушении
межклеточных связей в коре головного
мозга, например, после перерезки зрительного нерва и создания так называемого "мостика" из швановских
клеток путём имплантации периферического нерва
в область между клетками сетчатки глаза и четверохолмием. В этом случае аксоны перерезанного нерва
прорастают через мост, ветвятся и
образуют функционирующие синапсы в должных участках "своей"
мишени [6], c.
597.
Ещё одним примером индукционных
взаимодействий служит заживление ран и регенерация утерянных конечностей у
амфибий. У взрослых тритонов заживление раны начинается впервые двадцать четыре
часа после травмы. У аксолотлей на заживление раны уходит всего несколько
часов. У шпорцевых лягушек, даже у взрослых особей, при потере конечности, вырастает
новая [7].
В процессах Индукции активно участвует механизм генетической
памяти. Одно из важнейших свойств феномена Индукции – управления экспрессией генов
– производством белков, необходимых для развития организма. Это обстоятельство и ещё два
других фактора позволяют однозначно судить о приоритетной роли феномена
Индукции в процессах массового
тиражирования однотипных живых организмов в составе популяции.
Первый из двух факторов относится к свойству
ССП МО повышать эффективность информационного взаимодействия
с уменьшением расстояния [4]. Это свойство играет существенную роль в
предотвращении артефактов, обусловленных воздействием полей внешней среды.
Значимость второго
фактора обусловлена принадлежностью ССП МО
материальному
объекту – СПО МО является его атрибутом. Оно содержит
всю информацию о топологии материального
объекта, его элементном составе и
структуре вещества. Топология ССП данного объекта адекватна расположению
всех его структурных элементов на атомном уровне.
Из приведенных выше
экспериментальных результатов следует: специфика различных проявлений феномена
Индукции требует при их описании привлечения качественных – структурных и
топологических показателей. Известными физическими полями, удовлетворяющими
указанному условию, являются информационные Собственные спиновые поля всех
материальных объектов, включая поля кручения физического Вакуума [3].
Поскольку
воздействия слабых и сверхслабых полей внешней среды, в том числе полей
Физического Вакуума не могут непосредственно привести к разрушению полинуклеотидных
цепей ДНК, эти воздействия не могут изменить информационную матрицу, при
участии которой реализуется многократное воспроизводство данного вида живых организмов.
Для наглядности в
таблице 1 приведены основные характеристики
явлений взаимодействия МО и Индукции. Из таблицы 3 следует:
информационные процессы взаимодействия материальных объектов в явлении Индукции
подобны информационным процессам
взаимодействия в феномене Информационного взаимодействия ССП МО. В обоих
случаях результат взаимодействия определяется структурной составляющей
Собственных спиновых полей взаимодействующих объектов, несущих ин формацию об элементном составе их вещества, структуре
и топологии – форме, геометрических размерах объектов, расстоянии между ними. И
т.д.
Рассмотрение показателей в Таблице 1 приводит к заключению: феномены
Индукции и Информационного взаимодействия Собственных спиновых полей
материальных объектов – суть одно и то же явление, обнаруженное в разные эпохи
на различных объектах с применением различных методов. Индукция родилась в
эмбриологии, но ареал её деятельности гораздо шире: в её функции входит развитие растущих и обеспечение жизнедеятельности
взрослых организмов
Физическими факторами,
которыми оперирует феномен Индукции в процессах взаимодействия на микро-
уровне, являются те же основные факторы, что и в явлении Информационного
взаимодействия спиновых полей материальных объектов на макроуровне – структура
и топология.
Таблица 1. Сравнение основных
показателей в феноменах И ндукции и Явления
взаимодействия Собственных
спиновых полей материальных объектов
|
|
Явление Индукции |
Явление Взаимодействия ССП МО |
|
Год обнаружения |
2010 |
1901 |
|
Год раскрытия |
|
2011 |
|
Вид взаимодействия |
Информационный |
Информационный |
|
Природа
взаимодействия |
Спиновая |
Спиновая |
|
Категория основных исследуемых факторов |
Структура,
топология |
Структура, топология |
|
Материальные объекты |
Вещество,
сверхслабое постоянное магнитное поле соленоида (Н=10-5 э) |
Структуры биологических тканей,
сложные белковые молекулы, структурные кластеры в водных растворах
сверхнизкой концентрации |
|
Расстояния |
(10 – 2х10-12) м |
(10-1–1 0-7 – 10-1)
м |
|
Результаты
взаимодействия |
Изменение
состояния спинов спиновой системы веще-ства материального объекта, ведущее к
изменению его характеристик и свойств, и как следствие, к изменению физических
и физико-химичес-ких процессов, в которых объект участвовал |
Изменение состояния
Собственной спиновой системы биологической ткани, ведущее к изменению
в ней процессов
метаболизма и экспрессии генов, терапевтические эффекты гомеопатии |
Итак,
в явлении Индукции результат информационного взаимодействия определяется структурой
Собственных спиновых полей взаимодействующих объектов, несущей информацию об
элементном составе и топологических данных - форме, геометрических размерах и
расстоянии между объектами. На основании всего сказанного мы приходим к выводу:
явления Информационного взаимодействия спиновых полей материальных объектов и Индукции
– суть одно и то же явление, обнаруженное в разное время на различных объектах
и с применением различных методов.
По существу, весь объём
накопленных более чем за столетие, экспериментальных результатов, всесторонне
характеризующих явление Индукции, является
специфическим экспериментальным материалом обнаруженного в 2010 году
феномена Информационного взаимодействия ССП
МО. Специфика заключается,
во-первых, в живой природе исследуемых объектов; во-вторых, в малых дистанциях
между ними, в-третьих, в процессах, в которых явление Индукции себя проявляло.
Два сиамских близнеца – феномены
Индукции и Информационного взаимодействия собственных спиновых полей
материальных объектов впечатляют нетрадиционностью своего происхождения и
развития. Феномен, обнаруженный 110 лет
назад под именем "Индукция", родился в науке Эмбриология и до
сегодняшних дней развивался без понимания своих механизмов и своей природы.
Исторически явление Индукции не могло появиться в законченном виде даже на
полгода раньше. Только сегодня, когда явление Информационного взаимодействия
Собственных спиновых полей материальных объектов заняло своё место в физической
науке, стала понятной сущность явления Индукции и оно обрело свою
законченность.
Индукция – это физическое
явление, корнями своими ушедшее в биологию.
Индукция – это биологическое
явление физической природы. Его биологическая принадлежность была обнаружена в 1901
году. В следующие 112 лет феноменологическая разработка феномена проводилась с
целью определения его сущности, и только сегодня, когда явление Индукции
приобрело свою истинную природу, стало возможным обсуждение роли и значимости
физики в возникновении живых организмов. В отсутствии феномена Индукции,
планета Земля могла бы остаться без многоклеточных организмов – они просто не смогли бы образоваться.
Справедливости ради, добавим,
что одной физики совершенно не доста-
точно. Явление Индукции тесно взаимодействует
с явлением генетической памяти, которое не могло бы функционировать при
отсутствии рычагов управляющих экспрессией генов и генерацией белков,
необходимых для развития организма.
Литература
1. Бобров
А.В. Взаимодействие спиновых полей материальных объектов. Ч.1. Сознание и
физическая реальность. М.
"Фолиум", 2010, Т 15, № 7, с. 14-27.
2. Бобров А.В. Взаимодействие спиновых
полей материальных объектов. Ч.2,
Сознание и физическая реальность.
М., "Фолиум", 2010, Т
15, № 8 с. 9-26
5. Бобров А.В. Физическая природа
механизмов явления Индукции. Сознание и
физическая реальность. Фолиум, М., Том 16 № 8, 2011, с.40-63.
4.
Иванова Н.Н. Развернутый план-конспект урока биологии в 10-м классе по
теме "Онтогенез. Эмбриогенез". http://festival.1september.ru/articles/550303/
5. Албертс
Б., Брей Ц. и др. Молекулярная биология клетки. – М. "Мир", 1987, Т4,
с.111
6. Николс Дж. Г, А. Robert Martin и др. От Нейрона к мозгу. Изд. ЛКИ,
М.,
2008, с. 548-5536.
7. ресурс Интернета : http://www.zooclub.ru/amfib/l1/19.shtml