Биологические науки/ 5. Молекулярная биология
Ковалева1
Г. В., к.б.н. Ненашева1 В. В., к.б.н. Седова2 Е. С.,
проф., д.б.н. Тарантул1 В.
З.
1Институт
молекулярной генетики РАН, Москва, Россия
2ГУ НИИ
эпидемиологии и микробиологии имени Н.Ф. Гамалеи, Москва, Россия
Повышенная
экспрессия гена pub (TRIM-14) ассоциирована
как с ВИЧ-инфекцией, так и с поздними стадиями инфекции вируса гриппа
Ген pub, выявленный нами ранее как специфичный для
ВИЧ-ассоциированных лимфом (Nenasheva V. V. et al, 2005),
кодирует белок TRIM14, относящийся к семейству TRIM (Reymond
A. et al, 2001). Функции гена pub мало изучены. Известно, что его мышиный гомолог PUB связывается с транскрипционным фактором PU.1 и
оказывает существенное влияние на его активность (Hirose S. et al, 2003).
Кроме того, в ряде публикаций авторы наблюдали повышенную экспрессию гена pub при
различных инфекциях как вирусных, так и бактериальных. Так, например, было
обнаружено повышение экспрессии гена pub в периферической крови пациентов, инфицированных
следующими инфекционными агентами: вирусом гриппа типа A, грам-отрицательными (Escherichia coli) и
грам-положительными (Staphylococcus aureus, Staphylococcus pneumoniae)
бактериями, причем экспрессия pub была повышена
значительно у больных с гриппом по сравнению с больными с заболеваниями,
вызванными бактериями (Ramilo
O et al, 2007). Другие авторы
обнаружили повышение экспессии гена pub в печени шимпанзе, инфицированных вирусом гепатита В
на этапе виремии (Fakruddin J. M et al, 2007).
Исходя из данных литературы и результатов, полученных
нами ранее при анализе ВИЧ-ассоциированных лимфом (Nenasheva V. V. et al, 2005),
мы предположили возможное участие продукта гена pub в ответе организма на действие разных типов вирусов.
С целью проверки этой гипотезы в данной работе мы провели анализ экспрессии
гена pub и некоторых других генов в линии клеток карциномы
легких человека А549, трансфицированных вирусом гриппа птиц, адаптированным для
мышей (H5N2) (Smirnov Y.A et al, 2000).
Материалы и методы.
Клетки А549 культивировали в среде DMEM с 10% эмбриональной сывороткой КРС при 37оС
и 5%СО2 до достижения 70% конфлюэнтности. Вирус гриппа накапливали в 10-дневных куриных SPF
эмбрионах. Эмбрионы заражали в аллантоисную полость с множественностью 1000 ЭИД50
на эмбрион, инкубировали в течение 48 часов при 37°С, охлаждали в течение ночи
при 4°С и собирали вируссодержащую аллантоисную жидкость в стерильные пробирки.
Вируссодержащую аллантоисную жидкость хранили при 4оС или фасовали в
криопробирки по 1 мл и хранили при – 80оС. Инфекцию клеток проводили
в бессывороточной среде DMEM. После двух
часов инкубации вируса гриппа птиц, адаптированным для мышей A/Mallard
duck/PA/10218/84(H5N2) (Smirnov Y.A et al, 2000), с клетками среду снова
меняли на содержащую сыворотку. После 6, 21 и 24 часов инкубации из клеток
выделяли тотальную РНК с помощью реагента Тризола по методике производителя (Invitrogen, Carlsbad, CA, USA). Для
обратной транскрипции использовали набор Superscript III (Invitrogen) со случайной затравкой. Количественную ПЦР проводили
в iCycler thermal cycler (Bio-Rad Laboratories, Hercules, CA, USA) в
присутствии красителя SYBR Green и специфических праймеров для генов (pub: forward 5’-GCAGCAGCACATTGACAACA-3’, reverse 5’-TCCACGAGGCCCTTAAAGAA-3’;
set alpha: forward 5’- CGCCTCAAAAGAAGAAACCA-3’, reverse 5’-GCAGTGCCTCTTCATCTTCC-3’; set beta: forward 5’-AGCTCAACTCCAACCACGAC-3’, reverse 5’- GCAGTGCCTCTTCATCTTCC-3’, β-actin: forward 5’- TGCTTCTAGGCGGACTATGAC-3’, reverse 5’-AGAAGTGGGGTGGCTTTTAGG-3’). Условия амплификации: денатурация при 950С
10 мин, амплификация при 950С 15сек, 600С 30 сек, 720С
45 сек, 40 циклов. Относительные количества мРНК (гена pub, set alpha, set beta) были
посчитаны с помощью метода Ct,
рекомендованного производителем (Bio-Rad Laboratories,
Hercules, CA, USA), и нормализованы по количеству β-actin в тех же образцах.
Результаты исследования и обсуждение.
С помощью количественной ОТ-ПЦР мы проанализировали
изменение экспрессии гена pub, который был выявлен нами ранее как специфичный для
ВИЧ-ассоциированных лимфом, в клетках А549, инфицированных вирусом гриппа птиц,
адаптированным для мышей (H5N2), по сравнению с контрольными. Так же был взят для
исследования ген set, который, по нашим предыдущим данным (Nenasheva V. V.
et al.,
2005), повышенно экспрессировался как в спонтанных, так и ВИЧ-ассоциированных
лимфомах.
За 48-часовой период после заражения
вирусом линии А549 происходит лизис практически всех клеток. В течение первых
2-х часов после заражения идет синтез вирус-специфических полипептидов, в том
числе вирус-специфических компонентов, которые не включаются в вирион.
Приблизительно через 2 ч после заражения в клетках можно обнаружить антиген
нуклеопротеида. Почкующийся вирус может быть обнаружен на оболочках клеток
спустя приблизительно 5 ч после заражения. Примерно через 6 ч начинается выход
вируса в среду, и инфекционный вирус достигает максимальной концентрации в
среде через 7-8 ч после заражения.
На стадии 24 часов после заражения
наступает равновесие концентраций вирионов в среде и сохранившихся клетках с
разными этапами репликационного цикла. Поэтому для анализа были выбраны клетки
на стадии 6, 21 и 24 часа (период стабилизации клеток) после вирусной инфекции.
В результате данной работы была выявлено резкое
повышение экспрессии гена pub в клетках карциномы легких человека А549,
инфицированных вирусом гриппа, на стадии 24 часа, и понижение его экспрессии на
предыдущих стадиях инфекции (Рис.1). При этом экспрессия гена set (обеих форм
– альфа и бета) оставалась практически неизменной по сравнению с контролем на
стадии 24 часа при наличии понижения ранее (Рис. 2).
Подавление экспрессии всех генов на ранних
стадиях вирусной инфекции, возможно, объясняется активной работой вирусных генов,
сопровождаемой подавлением генов клетки-хозяина. Повышение экспрессии гена pub на более
поздних стадиях говорит о возможном участии гена в вирусном ответе в отличии от
гена set.
Таким образом, в результате нашей работы
мы обнаружили, что ген pub, кодирующий белок TRIM14, возможно, связан с вирусной инфекцией на поздних
этапах, что согласуется с данными литературы. Cемейство TRIM (tripartite motif) составляют белки, для которых характерно наличие так называемого TRIM (или RBBC) мотива, в
который могут входить три цинк-связывающих домена: RING (R), B-box 1 (B1) и B-box 2 (B2), а также coiled-coil (CC) регион (Reymond A et al,
2001). Так же у представителей TRIM семейства позвоночных (в частности, у
человека, мыши, цыпленка и лягушки) (Rhodes D. A.et al, 2005) встречается PRYSPRY домен.
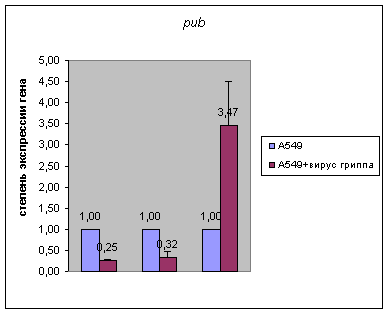
6ч
21ч 24ч
Рис. 1. Результаты количественной ОТ-ПЦР на мРНК из клетках карциномы легких человека А549, инфицированных вирусом гриппа, на разным стадиях после инфекции (6, 21 и 24 часа) для гена pub. В качестве контроля была использована РНК из неинфицированных клеток А549. Количество РНК было нормализовано по мРНК β-actin в тех же образцах.
Недавние исследования показали, что многие
TRIM белки вовлечены в широкий спектр биологических
процессов, включающих регуляцию транскрипции, контроль клеточной пролиферации и
дифференцировки, онкогенез и апоптоз, а также защиту клеток от вирусов (в
частности, от ВИЧ). На основании данных, полученных для нескольких TRIM белков (например, TRIM5alpha и TRIM19 (Towers G. J., 2007; Nisole S. et al, 2005)),
была предложена гипотеза о возможной роли PRYSPRY домена в иммунной защите от ретровирусов. Для разных
членов TRIM семейства цинк-связывающие домены В1 и В2 могут отличаться по
последовательности, однако интересно, что В boxes были обнаружены только у TRIM
белков и, видимо, являются их отличительной особенностью. Функции В boxes до
конца не выяснены, однако по последним данным они участвуют в защите клеток от
ВИЧ. Для TRIM5alpha было показано, что B box 2 влияет на узнавание С-концевым
доменом PRYSPRY вирусного капсида (Li
X. et al,
2007).
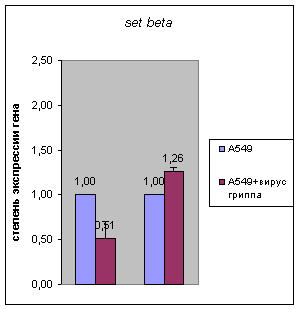
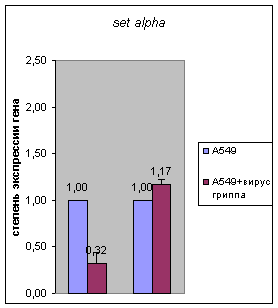
21ч 24ч 21ч 24ч
Рисунок 2.
Результаты количественной ОТ-ПЦР на мРНК из клетках карциномы легких человека
А549, инфицированных вирусом гриппа, на разным стадиях после инфекции (6, 21,
24 часа) для гена set beta и alpha. В
качестве контроля была использована РНК из неинфицированных клеток А549.
Количество РНК было нормализовано по мРНК β-actin в тех же образцах.
Таким образом, нами было обнаружен еще
один ген из семейства TRIM, который,
по-видимому, вовлечен в вирусный ответ клетки на действие вируса.
Литература:
1. Nenasheva V. V., Nikolaev
A. I., Martynenko A. V., Kaplanskaya I. B., Bodemer
W., Hunsmann
G., Tarantul
V. Z. Int J
Med
Sci. 2005. v.2, N. 4. p. 122-128.
2. Reymond A., Meroni G.,
Fantozzi A., Merla G., Cairo S., Luzi L., Riganelli D., Zanaria E., Messali S.,
Cainarca S., Guffanti A., Minucci S., Pellici P. G., Ballabio A. EMBO J. 2001. V. 20. P. 2140-2151.
3. Hirose S., Nishuzumi
H., Sakano H. Biocem. Biophs. Com. 2003.V. 311. P. 351-360.
4. Ramilo O, Allman W,
Chung W, Mejias A, Ardura M, Glaser C, Wittkowski KM, Piqueras B, Banchereau J,
Palucka AK, Chaussabel D. Blood. 2007. V. 109. N 5. P. 2066-2077.
5. Fakruddin JM,
Lempicki RA, Gorelick RJ, Yang J, Adelsberger JW, Garcia-Pineres AJ, Pinto LA,
Lane HC, Imamichi T. Blood. 2007. V. 109. N 5. P. 1841-1849.
6. Smirnov Y.A.,
Lipatov A.S., Van Beek R., Gitelman A.K., Osterhaus A.D., Claas E.C. Acta.
Virol. 2000. V. 44. P.1-8
7. Reymond A., Meroni G.,
Fantozzi A., Merla G., Cairo S., Luzi L., Riganelli D., Zanaria E., Messali S.,
Cainarca S., Guffanti A., Minucci S., Pellici P. G., Ballabio A. EMBO J. 2001. V. 20. P. 2140-2151
8. Rhodes D. A., de
Bono B., Trowsdale J. Immunology. 2005.
116. P. 411-417
9. Towers G. J. Retrovirology.
2007. 4:40.
10. Nisole S., Stoye J. P., Saib A. Nat. Rev. Microbiol. 2005. 3(10). P.
799-808
11. Li X., Song B., Xiang S. H., Sodroski J. Virology. 2007. 366. P.
234-244