Биологические науки/8.Физиология человека и
животных
Д. б.н. Хацаева Р.М.
Федеральное
государственное бюджетное учреждение науки Институт проблем экологии и
эволюции им. А.Н. Северцова РАН,
Москва, Россия
Этапы
развития обменных и пищеварительных процессов камер желудка сайгака в пренатальном онтогенезе
Изучение закономерностей развития обменных
и пищеварительных процессов
млекопитающих является одной их фундаментальных проблем эволюционной физиологии. Известно, что органы
пищеварения плода млекопитающих начинают функционировать уже в начале их закладки.
Питание плода осуществляется двумя путями: амниотрофным и гемотрофным, но их
развитие у всех видов происходит в разное время.
В настоящих исследованиях было прослежено
развитие обменных и пищеварительных функций камер желудка плодов сайгака (Saiga tatarica L., 1766) в возрасте от 45 дней
утробного развития до рождения, с подекадным чередованием. Для выявления
участия камер желудка сайгака в
обменных и пищеварительных процессах были применены гистологические,
гистохимические анализы на гликоген, гликопротеины, ферменты: липазу, щелочную
и кислую фосфатазы и биохимические на гликоген, пепсин и липазу.
Исследования показали, что камеры желудка у сайгака, с начала
своего формирования участвуют в углеводном обмене, о чем свидетельствует
наличие значительного количества
гликогена в слизистой оболочке всех камер желудка уже у 45-дневных плодов (рис.
1). В течение всего раннеплодного периода происходит депонирование гликогена в
сычуге, достигая максимума в 105 дней. С началом дифференцировки структур
слизистой оболочки сычуга начинается изменение локализации гликогена и
уменьшение его количества. К концу позднеплодного периода количество гликогена
снижается (рис.1).
В преджелудках ранних плодов сайгака
гликоген выявляется в пузырчатых клетках эпителия. Характер его
локализации говорит о расходовании на
процессы дифференцировки морфологических структур слизистых оболочек, начинающихся
раньше, чем в сычуге. Несмотря на расходование гликогена на процессы
дифференцировки, его количество в верхних
пузырчатых клетках на протяжении плодного периода увеличивается,
достигая максимума в 120 дней утробного развития (рис. 1), что является
свидетельством его использования для общих нужд плода, заменяя в какой-то
степени печень, гликогенобразовательная функция которой развивается в
позднеплодном периоде. К рождению, с повышением гликогендепонирующей функции печени, количество гликогена в преджелудках
значительно снижается (рис.1).
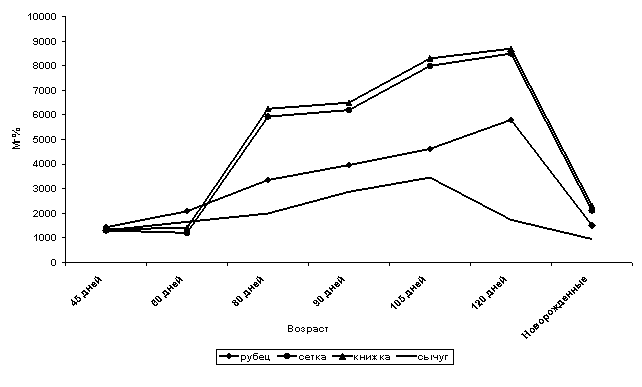 Рис. 1.
Активность гликогена в камерах плодов желудка сайгака.
Рис. 1.
Активность гликогена в камерах плодов желудка сайгака.
Об участии камер желудка в общем обмене
плода свидетельствует накопление
жидкого содержимого в полостях камер с момента их образования, до заглатывания амниотической
жидкости и начала секреторной деятельности желез сычуга. Обменные функции
осуществляются через межуточный обмен с помощью экскреции, направленной на
поддержание постоянства внутренней среды организма. В течение эмбрионального
периода экскреторная способность
осуществляется при активном участии эпителиальных клеток слизистой оболочки. В
раннеплодный период наблюдается образование большого количества вакуолей в эпителии, способствующих накоплению и
выведению веществ. По мере дифференцировки слизистой и развития на ее
поверхности гликокаликса, образование вакуолей затрудняется, и выделение
веществ в полость происходит через межклеточные пространства. К рождению экскреторная функция слизистой оболочки
камер желудка уменьшается, вакуоли в эпителии встречаются в основном в книжке.
Этому способствует и включение амниотрофного
питания в позднеплодный период путем заглатывания амниотической жидкости
(рис. 2).
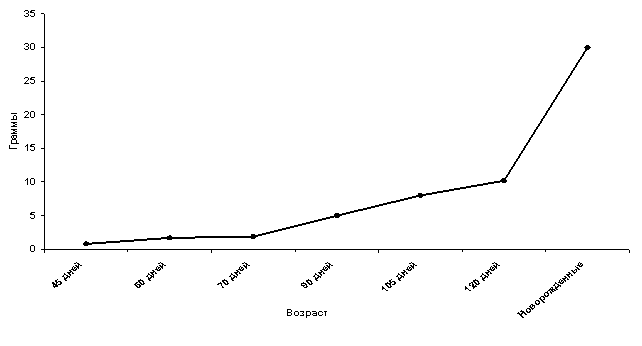
Рис. 2. Количество
содержимого в желудке у плодов сайгака
Вслед за обменными, включаются
пищеварительные функции желудка плода, о чем свидетельствует развитие желез в
сычуге. Первые признаки секреции гастронов у сайгака, как и у овец, появляются
в середине раннеплодного периода (60 дней). В состав их секрета в это время
входят: гликоген, нейтральные гликопротеины, следы липазы и пепсина. Позже
появляются кислые гликопротеины.
Активность пепсина в сычуге нарастает
постепенно до позднеплодного периода, и резко увеличивается к рождению (рис. 3).
Активность липазы повышается равномерно (рис. 3). Усиление активности ферментов
к рождению способствует более активному участию слизистой оболочки сычуга в
пищеварительных процессах. О транспорте веществ свидетельствует наличие в
эпителии камер желудка ферментов: липазы, щелочной и кислой фосфатаз, локализующихся
по клеточным границам эпителиального пласта, в эндотелии кровеносных сосудов и
нервных клетках.
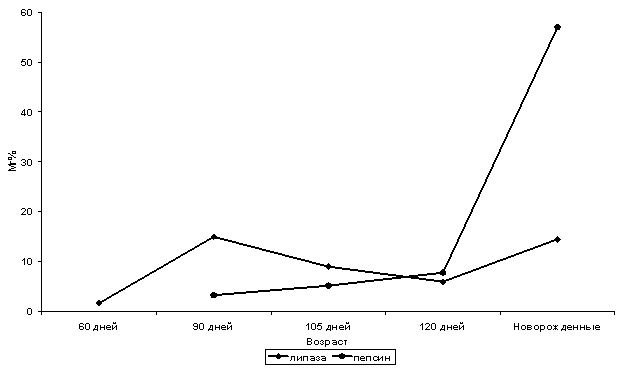
Рис.
3. Активность пепсина и липазы в сычуге плодов сайгака.
Таким образом, проведенные исследования
указывают на участие камер желудка
плодов сайгака, в обменных и пищеварительных процессах, обеспечивающих
питание плода амниотрофным и гемотрофным путями, благодаря их экскреторной
способности и ферментативной активности. Межуточный обмен в желудке плодов
сайгака, появляется с момента его закладки и уменьшается к рождению, а
пищеварительная способность возникает в середине раннеплодного периода,
увеличиваясь к рождению.
Ферментативная способность преджелудков
развивается интенсивно, формируя защитные свойства слизистой оболочки с
образованием гликокаликса. Это отражает у сайгаков большую значимость
преджелудков в обменных процессах и свидетельствует о яркой выраженности у них желудочно-кишечного типа пищеварения,
свойственного жвачным животным.