Романькова
К.М.1, Хромих Н. О.2, Лихолат Ю.В.1
Дніпропетровський національний університет імені Олеся Гончара
1Кафедра фізіології та інтродукції рослин
2 Науково-дослідний інститут
біології
ФУНКЦІОНУВАННЯ ГЛУТАТІОНОВОЇ СИСТЕМИ В ЛИСТКАХ КВІТКОВИХ РОСЛИН ЗА
УМОВ ГІПЕРТЕРМІЇ
При створенні
міських фітоценозів в екологічних умовах степового Придніпров’я, де висока
температура суттєво впливає на онтогенез рослин, необхідно здійснювати добір такого
складу газонних квіткових видів, який гарантує довговічність і декоративність фітоценозу.
Відомо, що внаслідок високотемпературного
стресу уповільнюються ріст і розвиток рослин, погіршуються декоративні якості, за
тривалої дії можливі незворотні реакції і загибель рослин [1; 2]. Проявом стресової дії гіпертермії на рослини
є посилене утворення у клітинах активних форм кисню (АФК) і порушення
окисно-відновної рівноваги, у відповідь на що активуються системи метаболічного
захисту, які забезпечують відновлення клітинного гомеостазу [2]. Важливий
механізм підтримання окисно-відновного стану у рослинних клітинах пов’язаний з
функціонуванням глутатіон-залежної системи, ефективність якої обумовлена загальним
пулом глутатіону, співвідношенням його відновленої
й окисненої форм (GSH/GSSG) та
активністю процесу відновлення глутатіону завдяки глутатіон-редуктазі (GR) [3; 4].
Мета роботи полягала
у виявленні закономірностей впливу гіпертермії на редокс-стан глутатіону та
активність глутатіон-редуктази у листках квіткових рослин. Об’єктами
дослідження були теплолюбні види цинія вузьколиста (Zinnia angustifolia L.) походженням з Центральної
Америки та гацанія блискуча (Gazania splendens hort.) з Південної Африки. Насіння рослин висівали у зволожений ґрунт у
вегетаційних посудинах, пророщували протягом 30 діб за кімнатної температури, проростки
піддавали дії високої температури (42˚ С) протягом 30 хвилин, 1 та 2
годин.
Установлено,
що за дії гіпертермії у листках рослин цинії вміст GSH несуттєво зростав, тоді як вміст GSSG помітно знижувався (рис. 1).
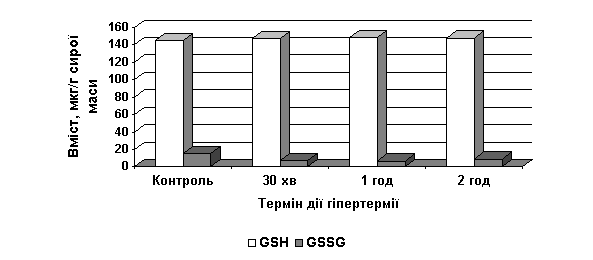
Рис. 1. Вплив гіпертермії (42˚ С) на вміст
відновленого та окисненого глутатіону (мкг/г сирої маси) у листках проростків Z. angustifolia
За
контрольних умов у листках цинії співвідношення GSH/GSSG становило 9,2; за впливу гіпертермії
(30 хв, 1 год, 2 год) воно зростало відповідно до 20,9, 22,0 та 16,4. Зменшення
вмісту GSSG вказувало на активацію
глутатіон-редуктази у листках цинії (таблиця).
Таблиця
Вплив гіпертермії (42˚ С) на
активність GR (нкат/хв•г
сирої маси)
у листках квіткових рослин
|
Термін дії |
Z. angustifolia |
G. splendens |
||||
|
M±m |
p |
% до К |
M±m |
p |
% до К |
|
|
Контроль |
75,041±0,072 |
- |
- |
87,693±0,150 |
- |
- |
|
30 хв |
76,049±0,251 |
0,0182 |
101,3 |
87,776±0,191 |
0,7494 |
100,1 |
|
1 год |
85,314±0,583 |
0,0006 |
113,7 |
88,986±0,218 |
0,0081 |
101,5 |
|
2 год |
92,895±0,382 |
0,0001 |
123,8 |
80,784±0,544 |
0,0003 |
92,1 |
Оскільки
активація GR у цинії
сягала найвищого рівня за тривалої дії гіпертермії, можна припустити, що
здатність до відновлення глутатіону більшою мірою визначає стійкість цинії до
температурного стресу, ніж зростання вмісту GSH, і таке припущення узгоджується
із закономірностями, встановленими для інших видів рослин [3].
У листках
гацанії вміст GSH достовірно
зростав протягом досліду, тоді як вміст GSSG зростав тільки через 30 хвилин,
надалі знижувався (рис. 2).
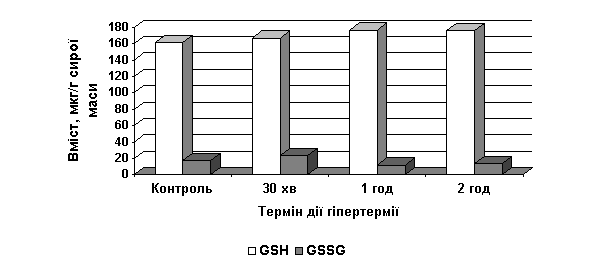
Рис. 2. Вплив гіпертермії
(42˚ С) на вміст відновленого та окисненого глутатіону (мкг/г сирої маси)
у листках проростків G. splendens
Співвідношення
GSH/GSSG у гацанії за контрольних умов
становило 9,0; за 30-хвилинного впливу гіпертермії знижувалось до 7,2, надалі
зростало до 14,8 і 13,0 (відповідно, за 1 і 2 год гіпертермії). При цьому
контрольний рівень активності GR у листках гацанії змінювався несуттєво (таблиця).
Таким чином,
за дії гіпертермії у листках гацанії більшою мірою було активовано процес біосинтезу
GSH, тоді як у листках цинії
– відновлення GSSG
завдяки активації GR. У
листках обох інтродукованих теплолюбних посухостійких видів виявлено зростання
співвідношення GSH/GSSG за дії гіпертермії, що
узгоджується з оцінкою такої реакції глутатіонової системи як показника
стійкості рослин до інших типів стресорів [4].
Література:
1. Almeselmani M., Deshmukh P.S., Sairam R.K., Kushwaha S.R., Singh T.P. //
Plant Science. – 2006. – V. 171, N 3. – P. 382-388.
2. Ma Y.-H., Ma F.-W., Zhang J.-K., Li M.-J., Wang Y.-H., Liang D. // Plant Science. –
2008. – V. 175, N6. – P. 761-766.
3.
Noctor G., Arisi A.C.M., Jounanin L., Kunert K.J., Rennenberg H., Foyer C.H. // J. Exp. Bot. 1998.
– V. 49, N 321. P. 623–647.
4. Schmieden U.,
Schneider S., Wild A. // Env. Pollution. – 1993. – V. 82, N 3. – P. 235-244.