Взаимосвязь между содержанием
катехоламинов в мозжечке и некоторых структурах эмоциогенной
лимбико-неокортикальной системы мозга у крыс с доминантным и субмиссивным
типами поведения
Попова Л.Д., Васильева И.М.
Харьковский
национальный медицинский университет
Кризисные
процессы, происходящие в современном обществе, негативно влияют на психологию
людей, вызывая тревожность, напряжение, жестокость и насилие (1).
Исследование
особенностей нейрогуморального статуса у животных, склонных к доминантному или
субмиссивному типу поведения имеет большое значение для понимания
нейробиологических механизмов развития агрессии и депрессии у человека. Нейрофизиологические
и биохимические исследования нейромедиаторного обеспечения формирования
агрессивных и депрессивных состояний, в основном, сфокусированы на структурах
эмоциогенной лимбико-неокортикальной системы мозга. В то же время, роль
мозжечка в этих процессах практически не изучена, хотя мозжечок, который
является важной структурой регуляции
локомоторной функции организма, очевидно, имеет отношение к реализации
агрессивного поведения. Кроме того, в последние годы появились работы, согласно
которым мозжечок принимает участие в регуляции высших когнитивных (2) и
психоэмоциональных функций (3). Некоторые авторы считают мозжечок незаменимым
узлом в нейрональных цепях, поддерживающих высшие уровни поведения (4).
В связи
с этим, целью нашей работы было изучение взаимосвязей между содержанием катехоламинов, играющих важную роль в
развитии депрессивных и агрессивных состояний, в мозжечке и фронтальной
коре/гиппокампе.
Материалы и методы. Работа выполнена на
26 белых лабораторных крысах-самцах
молодого репродуктивного периода (6 мес.), которые содержались в стандартных
условиях вивария. Манипуляции на
животных проводились в соответствии с положениями «Общих этических принципов
экспериментов на животных», одобренных Первым Национальным конгрессом по
биоэтике (Киев, 2001). Для разделения животных на группы с альтернативными
типами поведения была использована модель эмоциогенного стресса «Сенсорный
контакт» с некоторыми модификациями (5, 6). Согласно этой модели, животные в
течение 5 суток находились в условиях индивидуального содержания с целью
предупреждения эффекта группового взаимодействия. Потом крыс содержали 2 дня в
экспериментальных клетках, разделенных на два равных отсека прозрачной
перегородкой с отверстиями, что обеспечивало условия сенсорного контакта.
Тестирование типа поведения проводили
через 2 дня после адаптации животных к новым условиям содержания и сенсорного
знакомства. В период тестирования перегородку убирали на 10 минут. Тестирование
проводили в течение 10 дней во второй половине дня (14.00-16.00).
Содержание
катехоламинов в структурах головного мозга определяли флуометрическим
микрометодом (7). Для экстракции моноаминов 5мг ткани головного мозга
гомогенизировали в 1.0 мл смеси НСl-бутанола. После центрифугирования в течение
20 минут при 2000g надосадочную фракцию переносили в пробирку, содержащую 2.0 мл
гептана и 0.25 мл 0.1 М НСl. В течение 10 минут пробирку встряхивали, потом
центрифугировали в течение 10 минут при 2000g. После этого водную фазу
использовали для определения норадреналина (НА) и дофамина (ДА). После
окисления катехоламинов измеряли флуоресценцию НА на спектрофлуориметре «Hitachi» (длины волн возбуждения и люминесценции составляли,
соответственно, для НА – 395 и 485, для ДА – 330 и 375нм).
Статистическую
обработку полученных результатов проводили методами непараметрической
статистики при помощи пакета общего назначения «Statistica 6.0». Для выявления зависимости показателей от группы
использовали непараметрические аналоги дисперсионного анализа – критерии Крускала-Уоллиса
и медианный тест. Для попарного сравнения групп использовали критерий Манна-Уитни и корреляционный анализ по
Спирмену.
Результаты исследования
Согласно
полученным результатам, во всех изученных структурах содержание норадреналина у
субмиссивных и доминантных самцов достоверно отличались от содержания данного
моноамина у уравновешенных животных. При этом у субмиссивных самцов наблюдалось
повышение, а у доминантных -понижение данного
показателя (табл.1).
Таблица 1
Содержание
норадреналина (нмоль/г ткани) в структурах головного мозга крыс с
альтернативными типами поведения
Тип поведения
|
Фронтальная кора |
Гиппокамп |
Мозжечок |
||||
|
Медиана Ме |
Квартили 25%; 75% |
Медиана Ме |
Квартили 25%; 75% |
Медиана Ме |
Квартили 25%; 75% |
||
Субмиссивный
|
7.88*,** |
5.81; 9.21 |
8.92*,** |
6.27; 9.48 |
4,51*,** |
4,05; 4,81 |
|
|
Уравновешенный |
4.65 |
4.65; 5.03 |
5.79 |
4.34; 5.79 |
2,79 |
2,17; 3,11 |
|
|
Доминантный |
2.32* |
2.32; 3.48 |
3.37* |
2.89; 4.34 |
2,17* |
1,86; 2,76 |
|
Примечание: * -
р<0.05 по сравнению с самцами с уравновешенным типом поведения; ** - р<0.05 по сравнению с самцами с
доминантным типом поведения.
Полученные
нами результаты согласуются с большинством литературных данных, согласно
которым депрессия чаще ассоциируется с избыточной активностью НА-ергической
системы (8). По данным литературы, стресс сопровождается освобождением НА в
большинстве структурах головного мозга (9). Поскольку гиппокамп опосредованно
вовлекается в медиацию начала ответа на стресс, а также способствует контролю
продолжительности ответа на стресс (10), повышенное содержание НА в этих
структурах у субмиссивных животных будет способствовать гиперактивации
гипоталамо-гипофизарно-адреналовой системы, развитию депрессивных состояний и
импульсивной агрессии, которая может проявляться на фоне депрессии. Фронтальная
кора управляет когнитивными функциями, оказывает ингибирующее влияние на
агрессию и жестокость (11). Атрофия фронтальной коры приводит к патологической
жестокости, сопровождается уменьшением чувствительности к боли. Возможно, что уменьшение
содержания НА в этой структуре у доминантных
самцов имеет отношение к проявлению адаптивной агрессии в этой группе
животных.
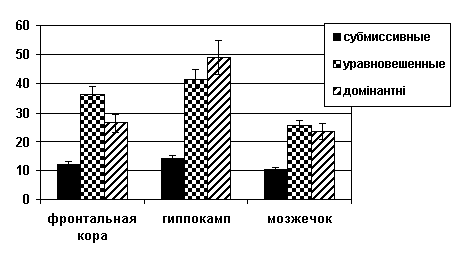 При изучении содержания ДА обнаружено
его достоверное снижение во всех исследуемых структурах у субмиссивных самцов
по сравнению как с уравновешенными, так и доминантных крысами. Разница в
содержании ДА между доминантными и уравновешенными животными наблюдается только
во фронтальной коре (рис. 1).
При изучении содержания ДА обнаружено
его достоверное снижение во всех исследуемых структурах у субмиссивных самцов
по сравнению как с уравновешенными, так и доминантных крысами. Разница в
содержании ДА между доминантными и уравновешенными животными наблюдается только
во фронтальной коре (рис. 1).
![]()
![]()
![]()
![]()
Рисунок 1. Содержание
дофамина (нмоль/г ткани) в структурах головного мозга крыс с альтернативными
типами поведения.
Примечание: * - р<0.05 по сравнению с самцами с уравновешенным типом поведения; ** - р<0.05 по сравнению с самцами с доминантным типом поведения.
Дофаминергические мезолимбические и
мезокортиколимбические пути имеют проекции от вентрального мезенцефалона к
лимбическим и кортикальным структурам, регулирующим когнитивные и эмоциональные
функции (12). Полученные результаты согласуются с мнением Olanow C.W., Kollor
W.C. (2001) о том,
что основное значение в патогенезе депрессии, в частности у больных
паркинсонизмом, может иметь поражение мезокортиколимбических дофаминергической
и серотонинергической систем, модулирующих состояние фронтостриарных кругов и
лимбических структур (13).
Анализ корреляционных связей между
содержанием катехоламинов в мозжечке и
структурах эмоциогенно-неокортикальной системы выявил существование у доминантных
самцов тесной положительной связи между содержанием НА в мозжечке и фронтальной
коре (+0.746, р<0.05), содержанием НА в мозжечке и гиппокампе (+0.704, р<0.05).
Кроме этого, как у доминантных, так и субмиссивных самцов обнаружена высокая
положительная корреляционная связь между содержанием ДА в мозжечке и
фронтальной коре (субмиссивные: + 0.881, р<0.05; доминантные: + 0.779,
р<0.05), в мозжечке и гиппокампе (субмиссивные: + 0.887, р<0.05; доминантные:
+ 0.683, р<0.05).
Выводы. Принимая во внимание нейронные связи мозжечка с корой
мозга и структурами лимбической системы, однонаправленность изменений в содержании
катехоламинов в мозжечке, фронтальной коре и гиппокампе, существование
корреляционных связей между содержанием катехоламинов в мозжечке и структурах
эмоциогенной лимбико-неокортикальной системы, можно предположить, что мозжечок
вовлекается в формирование доминантного/субмиссивного типов поведения, в
развитие агрессивных и депрессивных состояний.
Литература
|
1. |
Ho H.P. The serotonin reuptake inhibitor fluoxetine reduces sex
steroid – related aggression in female rats: an animal model of premenstrual
irritability? / H.P. Ho, M. Olsson, L. Westberg [et al.]//
Neuropsycho-pharmacology. – 2001. – Vol. 24, N5. – P.
502-5011. |
|
2. |
Steinlin M. Cerebellar disorders in
childhood: cognitive problems / М. Steinlin // Cerebellum. -
2008. – Vol. 7, N 4. – P. 607-610. |
|
3. |
Ito M. Long-term depression. / M.
Ito // Annu Rev Neurosci. – 1989. – N 12. – P. 85-102. |
|
4. |
Levisohn
L. Neuropsychological consequences of cerebellar tumour resection in
children
Cerebellar cognitive affective syndrome in a paediatric population /L. Levisohn,
A.
Cronin-Golomb, J.D.
Schmahmann //Brain. - (2000. – Vol.
123, N 5. – P. 1041-1050. |
|
5. |
Kudryavtseva N.N.
The sensory contact model for the study of aggressive and submissive
behaviors male mice/ N.N. Kudryavtseva //Aggres. Behav. – 1991. – Vol. 17,
№5. - P.285-291. |
|
6. |
Патент на корисну модель № 55046 МПК А61В5/16
«Спосіб визначення типу поведінки експериментальних гризунів», Попова Л.Д.,
Васильєва І.М. Харківський національний медичний університет. Заявка №U201002821від 12.03.2010.Опубл.: 10.12.2010 Бюл.№23 |
|
7. |
Schlumpf M. A
Fluorometric micromethod for the simultaneous determination of serotonin,
noradrenaline and dopamine in milligram amounts of brain tissue / M.
Schlumpf, W. Lichtensteiger, H. Langemann [et al.] //Biochemical
Pharmacology. – 1974. – Vol. 23, N 17. – P. 2437-2446 |
|
8. |
Ressler K.J. Role
of serotonergic and noradrenergic systems in the pathophysiology of
depression and anxiety disorders. / K.J. Ressler, C.B. Nemeroff // Depress
Anxiety. – 2000.
– Vol. 12, Suppl 1. – P. 2-19. |
|
9. |
Tanaka M.
Emotional stress and characteristics of brain noradrenaline release in the
rat. / M. Tanaka // Ind Health. – 1999. – Vol. 37, N2. P. 143-156. |
|
10. |
Herman J.P. Role of the ventral subiculum in stress integration./ J.P.
Herman, N.K. Mueller // Behav
Brain Res. – 2006. – N174. – P. 215–224. |
|
11. |
Ferris C.F.
Vasopressin and serotonin interactions in the control of agonistic behavior.
/C.F. Ferris, Y. Delville // Psychoneuroendocrinology. 1994. – Vol. 19, N5-7.
– P. 593-601. |
|
12. |
Liberman A.
Depression in Parkinson's disease: a rewiew / A. Liberman // Acta Neurol.
Scand. – 2006. – Vol. 113. – P. 1-8 |
|
13. |
Olanow C.W. An
algorithm (decision tree) for the management of Parkinson's Disease:
treatment guidelines / C.W. Olanow, W.C. Koller // Neurology 2001. – Vol. 50,
Suppl. 3. – P. 1-63 |