ИННОВАЦИОННЫЕ ТЕХНОЛОГИИ В СЕЛЕКЦИИ
КРУПНОГО РОГАТОГО СКОТА
Фенченко Н.Г., профессор, доктор
с.-х. наук, зав. лаб. селекции и воспроизводства мясного скотоводства;
Хайруллина Н.И., доктор с.-х. наук,
вед. науч. сотр. лаб. селекции и воспроизводства мясного скотоводства;
Галимов Р.Ф., канд. с.-х. наук,
младш. науч. сотр. лаб. селекции и воспроизводства мясного скотоводства
Шайхутдинова А.З., зооинженер;
Ильина Н.В., аспирант лаб. селекции
и воспроизводства мясного скотоводства;
* Гафарова Ф.М. кандидат с.-х. наук
ФГБОУ ВПО Башкирский государственный аграрный университет
450059 Республика Башкортостан, г.
Уфа, Рихарда Зорге, 19, ГНУ Научно-исследовательский институт сельского
хозяйства Российской академии сельскохозяйственных наук, bagri@ufanet.ru. 8(347) 223-09-56.
Комплексное изучение и
внедрение в практику селекционно-генетических методов позволит создать
высокопродуктивные чистопородные и помесные стада крупного рогатого скота
адаптированного к разведению в условиях резко-континентального климата,
обеспечивающих интенсивность роста, развития молодняка и повышение молочной
продуктивности взрослого поголовья племенных стад крупного рогатого скота
чёрно-пёстрой и голштинской пород.
Ключевые слова: порода; линия;
селекция; микросателлиты; ДНК; продуктивность; аллель; генотип; полиморфизм;
локус; маркеры; генетический потенциал; генеалогия.
Comprehensive
study and implementation in practice of selection and genetic techniques will
create high-yield pure-bred and cross-bred herds of cattle adapted to breeding
in a sharply continental climate, providing the intensity of growth,
development and improvement of young dairy cattle breeding productivity of
adult cattle herds Black and White and Holstein breeds.
Keywords:
breed; line; selection; microsatellites; DNA; productivity; allele; genotype;
polymorphism; locus; markers; genetic potential; genealogy.
N.G.
Fenchenko, N.I. Khairullina, R.F. Galimov, A.Z. Shaikhutdinova, N.V. Ilina,
F.M.Gafarova
l. Введение
В современных условиях для объективной оценки разнообразия
пород сельскохозяйственных животных используемых в животноводческих отраслях
поддерживается их разведение в чистоте с учётом генеалогической структуры.
Установлено, что для этой цели в мире максимально
используются методы ДНК-биотехнологии основанные на анализе материалов
полиморфизма. Среди них особая роль отводится приоритетным методам
микросателлитного анализа ДНК (1-4).
Отбор животных позволяет учитывать новые качества, такие как
приспособленность к условиям содержания, устойчивость против различных
заболеваний, которые должны быть положены в основу селекции, основанной на
изучении изменчивости, наследуемости и взаимозависимости признаков, существенно
меняющиеся из поколения в поколение.
Следовательно, необходима система постоянного контроля
изменения генетической ситуации в данном массиве животных с тем, чтобы вносить
необходимые коррективы в программу селекционной работы с ним.
ll. Методы исследования
Научно-исследовательская работа проводилась на поголовье
чёрно-пёстрой породы племенных племрепродукторах ОАО «Зирганская МТС»
Мелеузовского района, СПК им. Салавата
и ОАО Агрофирма «Правда» Стерлибашевского района.
Объектом исследований служили ремонтные телки, коровы,
быки-производители черно-пестрой породы Уральского отродья, разводимые в зоне
Южного Урала, которые содержались при оптимальных условиях кормления и
содержания в соответствии с зоотехническими и зоогигиеническими требованиями.
Рационы для кормления животных составлялись с учетом норм кормления ВИЖа из
кормов собственного производства.
Животные в группы подбирались с учетом времени рождения,
возраста, даты отела, продуктивности матерей, линейного происхождения,
кровности по голштинской породе коровы черно-пестрой породы Уральского отродья
датской, немецкой и голштинской пород в количестве 200 голов коров.
Анализу подлежали экспериментальные данные, полученные в
научно-хозяйственных и производственно-исследовательских опытах, при
использовании данных зоотехнического, племенного учета продуктивности скота
черно-пестрой породы различного экогенеза, материалы бонитировки животных, акты
контрольных доек, взвешивания животных, журналов выращивания ремонтного молодняка,
искусственного осеменения и отелов коров, карточки племенных коров 2-Мол,
карточки племенных быков 1-Мол, ГКПЖ и каталогов быков-производителей,
оцененных по качеству потомства.
При изучении генотипического различия чёрно-пёстрого скота
использовали метод зоотехнического анализа на основе программы СЕЛЕКС и
молекулярно-генетический (микросателлитный).
Использование генетической структуры племенного стада и
материалов генетической полимеризации микросателлитов обеспечило возможность
прогнозировать продуктивность, идентифицировать биологический индивид и выявить
родство и различие линейной принадлежности маточного поголовья крупного
рогатого скота чёрно-пёстрой породы.
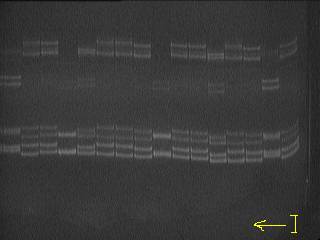
Рис. 1. Микросателлитные локусы геномного ДНК выделенные из
крови чёрно-пёстрой породы крупного рогатого скота Уральского отродья
Показана роль ДНК-технологий при оценке племенных качеств
быков производителей с учётом реализации их генетического потенциала в
продуктивных качеств дочерей в зависимости от линейной принадлежности.
На основе данных зоотехнического и племенного учёта была
определена генеалогическая структура популяции чёрно-пёстрого крупного рогатого
скота с установлением породной и линейной принадлежности положенная в основу
перспективного плана селекционно-племенной работы. По результатам ежегодной
бонитировки были сформированы группы маточного поголовья потомства семи
быков-производителей относящихся к четырём линиям на которых и проводили
изучение их генетического полиморфизма.
lll. Результаты исследований
В результате
исследований установили, что по идентификации образцов крови из 11 определяемых
локусов у крупного рогатого скота было выявлено наличие у подопытных животных
шести ВМ 1824, ВМ 2113, ЕТН 10, ЕТН 35, SPS 115 и TGLA с конкретным их распределением
(таблица 1). Определенные микросателлитные маркеры показывают на существенное
различие по частоте встречаемости отдельных аллелей микросателлитных локусов.
В частности по локусу ВМ 1824 с размером маркера 178-192
п.н. и первым аллельным методом микросателлитных локусов минимальные показатели
повторяемости составили 045-064 у потомков быков-производителей Мудрого 2819
линии Атлета 4, Кадра 1386, Лобнера 10013 линии А.Адема 30586 черно-пестрой и
М.Чифтейна 95679 голштинской пород.
Относительно высокой повторяемостью 1,63 аллелей
микросателлитных локусов характеризовалось потомство быка Янаула 4909 линии
Атлета 4.
Несколько ниже показатели 0,99-1,18 прослеживались у дочерей
быков-производителей Символа 476 и Банкира 109 линии А.Адема 30586, тогда как у
их сверстниц быка Бонуса 36 данный показатель был несколько ниже и составил
0,73. В этом отношении выделяются животные второй аллельной группы
микросателлитов локуса ВМ1824 с размером маркера 178-192, где частота варианты
почти по всем быкам-производителям была минимальной и составила 0,18-0,36 кроме
Банкира 109 и Символа 476 при 0,73. При этом в данном локусе выявлено всего
лишь пять аллельных вариантов при восьми и девяти у локусов ЕТН 995 и TGLA 122 с размером маркера 131-159 и
131-181.
Между тем, практически отсутствовали четвертая или пятая у
потомков всех быков-производителей первого локуса кроме Банкира 109 с частотой
встречаемости 0,09-0,36.
Практически не обнаружены микросателлиты по локусу TGLA 122 у дочерей быков-производителей
с седьмой по девятую аллельному варианту Банкира 109, третьей Символа 476, второй,
третьей и пятой Лобнера 10013, Кадра 1386, от седьмой по девятую у остальных, а
также первой у Янаула 4909, пятой Мудрого 2819 и третьей Бонуса 36.
Тем не менее, выделяются животные с локусом ЕТН 10, где у
всех их независимо от быка-производителя отсутствует первый и пятый вариант
аллельных микросателлитов, тогда как по локусу ВМ 2113 с размером маркера
129-143 п.н. повторяемость аллельных вариантов были представлены всего лишь с
первой по четвертый микросателлитных локусов. Однако частота встречаемости
аллельных микросателлитов по локусу ЕТН 995 была минимальной 0,09-0,36 за
исключением дочерей быков-производителей Банкира 109, Янаула 4909, Символа 476
и Кадра 1386 при 0,64-1,54.
Между тем по локусу ВМ 2113 с размером маркера 123-143
показатели повторяемости при минимальном количестве аллельных вариантов
составило 0,45-1,36, тогда как по четвертой аллели локуса TGLA 122 она была максимальной 1,18-1,63
потомства быков Банкира 109 и Символа 476.
С седьмой по девятую аллель прослеживались минимальные ее
показатели 0,09-0,18.
Табл. 1. Частота встречаемости аллелей микросателлитных
локусов дочерей быков-производителей разных линий черно-пестрой породы
|
Локус,
размер маркера п.н. |
Алле-льные
вариан ты
микро-сател- литных
локу-сов |
Кличка
и инвентарный № быка-производителя |
||||||
|
Банкир
109 |
Символ
476 |
Кадр
1386 |
Лобнер
10013 |
Янаул
4909 |
Мудрый
2819 |
Бонус
36 |
||
|
ВМ
1824 178-192 |
1 |
1,18 |
0,99 |
0,64 |
0,64 |
1,63 |
0,45 |
0,73 |
|
2 |
0,73 |
0,73 |
0,27 |
0,27 |
0,36 |
0,18 |
0,18 |
|
|
3 |
1,54 |
1,54 |
1,21 |
0,54 |
1,72 |
0,54 |
0,64 |
|
|
4 |
0,36 |
0,18 |
- |
- |
- |
0,09 |
- |
|
|
5 |
0,09 |
- |
0,27 |
- |
0,18 |
- |
0,09 |
|
|
ВМ
2113 123-143 |
1 |
1,36 |
1,18 |
- |
0,45 |
0,99 |
0,45 |
- |
|
2 |
0,18 |
0,09 |
- |
0,45 |
0,99 |
0,45 |
0,27 |
|
|
3 |
0,45 |
0,18 |
0,54 |
0,27 |
0,54 |
0,27 |
- |
|
|
4 |
0,99 |
1,18 |
0,54 |
0,36 |
0,64 |
0,36 |
- |
|
|
5 |
- |
0,64 |
0,09 |
- |
0,18 |
- |
0,27 |
|
|
ЕТН
10 207-231 |
1 |
0,18 |
0,18 |
0,18 |
0,27 |
0,45 |
- |
0,27 |
|
2 |
1,54 |
1,81 |
0,91 |
0,64 |
1,81 |
0,64 |
0,82 |
|
|
3 |
0,27 |
0,09 |
0,09 |
- |
- |
0,09 |
0,09 |
|
|
4 |
- |
- |
0,09 |
0,09 |
- |
- |
- |
|
|
5 |
1,54 |
1,54 |
0,82 |
0,45 |
1,27 |
0,54 |
0,45 |
|
|
ЕТН
225 131-159 |
1 |
- |
0,54 |
0,64 |
0,36 |
1,09 |
0,27 |
0,82 |
|
2 |
0,36 |
- |
0,09 |
0,18 |
- |
0,27 |
- |
|
|
3 |
0,36 |
0,27 |
0,27 |
0,36 |
0,45 |
0,27 |
0,45 |
|
|
4 |
1,54 |
1,27 |
0,82 |
0,36 |
1,18 |
0,36 |
0,27 |
|
|
5 |
0,27 |
0,36 |
0,09 |
0,09 |
- |
- |
- |
|
|
6 |
0,09 |
- |
- |
- |
0,27 |
- |
0,09 |
|
|
7 |
0,73 |
0,64 |
0,27 |
0,09 |
- |
- |
- |
|
|
8 |
- |
0,09 |
- |
- |
0,09 |
0,09 |
- |
|
|
SPS 115 234-258 |
1 |
0,82 |
0,27 |
0,09 |
0,18 |
0,91 |
0,36 |
0,18 |
|
2 |
1,09 |
1,18 |
0,64 |
0,54 |
- |
- |
- |
|
|
3 |
0,54 |
0,91 |
0,36 |
0,36 |
1,09 |
0,64 |
0,36 |
|
|
4 |
1,09 |
0,64 |
0,18 |
0,27 |
0,91 |
0,27 |
0,64 |
|
|
5 |
0,18 |
0,64 |
0,45 |
0,09 |
0,73 |
- |
0,09 |
|
|
6 |
- |
- |
0,27 |
- |
- |
- |
0,09 |
|
|
7 |
- |
- |
- |
- |
0,18 |
- |
- |
|
|
TGLA 122 137-181 |
1 |
0,91 |
1,09 |
0,54 |
0,54 |
- |
0,54 |
0,27 |
|
2 |
0,54 |
0,18 |
- |
- |
0,91 |
0,09 |
- |
|
|
3 |
0,09 |
- |
- |
- |
0,09 |
- |
0,27 |
|
|
4 |
1,63 |
1,18 |
0,54 |
0,54 |
1,36 |
0,54 |
0,18 |
|
|
5 |
0,09 |
0,09 |
0,09 |
- |
0,18 |
- |
0,09 |
|
|
6 |
0,54 |
0,27 |
0,36 |
0,09 |
0,18 |
0,09 |
0,09 |
|
|
7 |
- |
0,09 |
0,09 |
0,18 |
- |
- |
0,09 |
|
|
8 |
- |
0,18 |
0,18 |
0,09 |
- |
- |
- |
|
|
9 |
- |
0,09 |
0,09 |
0,09 |
- |
- |
- |
|
Анализ генеалогической структуры стада показал, что
прослеживается определенная тенденция определяющая взаимосвязь между частотой
встречаемости определенных аллельных вариантов микросателлитных локусов ДНК и
показателями продуктивности животных с учетом их линейной принадлежности
(таблица 2).
Из приведенных данных следует, что по молочной
продуктивности выделяются первотелки быков-производителей Банкира 109, Символа
476 и Кадра 1386 линии А.Адема 30586 с удоем за 305 дней лактации от 6274±134кг
до 6943±247кг и живой массой 500-692кг.
Между тем у них же был максимальный коэффициент
повторяемости аллельных вариантов микросателлитных локусов от 24 до 65 или
2,18-5,89% с первой по шестую аллель независимо от их нахождения в локусе.
Несколько уступали им представители линии Р.Соверинга 198998
и Атлета 4, которые являлись потомками быков-производителей Бонуса 36 и Янаула
4909 с удоем 5133-5679кг и живой массой 521-572кг с первой 22-63 или 2,63-5,72%
по четвертую аллель микросателлитных локусов. Тем не менее по МДЖ существенных
различий практически не установлено 3,63-3,78% исключая потомков быков Символа
476 и Лобнера 10013 от 4,11-4,18.
При низкой продуктивности животных, величина генетического
тренда по удою в сравнении с высокопродуктивными сверстницами, в 2,9 раза ниже
(таблица 3).
Незначительные различия абсолютного прироста генетического
потенциала обусловлены генеалогической и генотипической схожестью стад,
следовательно, изначально животные имели сходный генетический потенциал
продуктивности. В стадах с удоем до 4000 кг ежегодный реализованный прирост
генетического потенциала 16,2% от абсолютного прироста генетического потенциала
и 46,4% от фенотипического роста продуктивности. Это свидетельствует о
недостаточной подготовке этих хозяйств к разведению высокопродуктивного скота.
Так как сложившиеся недостаточно комфортные условия среды не только не
позволяют проявиться в полной мере генетическому потенциалу, но и ведут к
элиминации генов голштинской породы, как более требовательных.
lV. Выводы
Впервые разработаны и использованы селекционно–генетические
приемы управления селекционным процессом для создания высокопродуктивных
чистопородных племенных стад крупного рогатого скота, перспективных линий и
пород, адаптированных к разведению в условиях резко-континентального климата,
обуславливающих выявление дополнительных резервов повышения живой массы и
молочной продуктивности, за счет увеличения ввода в основное стадо
высокоценного племенного молодняка крупного рогатого скота.
Таким образом, при максимальном использовании в селекции
ДНК-технологий появляется возможность выявления дополнительных резервов
увеличения производства молока и улучшения его качества за счёт интенсивного
использования высокоценных быков–производителей перспективных линий, закладки
новых семейств обеспечивающих сохранение генофонда чёрно-пёстрой породы
крупного рогатого скота Уральского отродья.
ЛИТЕРАТУРА
1. Aberle K.S., Hamann H., Drogemuller C., e.a.
// Genetic diversity in German draught horse breed compared
with a group of primitive, riding and wild horses by means of microsatellite
DNA markers. Animal Genetics. -2004.-N. 35.-P. 270-277.
2. Luis
C, J u r a s R., О о m M.M., e.a. // Genetic diversity and relationships of Portuguese and other horse
breeds based on protein and microsatellite loci variation. Animal Genetics. –
2007.- N.38.-P. 20-27.
3. Bjernstad, G. // Breed demarcation and
potential for breed identification of horses assessed by molecular markers /
G.Bjornstad, K. H.Roed // 51st Annual Meeting of the European
Association for Animal Production.-Hague (Netherlands).- 2000.-N.6.-P. 354-355.
4. Williams J.L, Haley C.S. // Discriminating
among cattle breeds using genetic markers. Heredity.-1999.-N.82.-P.613-619.
5. Bowcock, A.M. // High resolution of human
evolutionary trees with polymorphic microsatellites / A.M.Bowcock ef al. /7
Nature. -1994.-V.368.-P. 455-457.
6. Фенченко Н.Г., Хайруллина Н. И., Сиразетдинов Ф.Х. История
создания и генеалогия чёрно-пёстрой породы крупного рогатого скота. Уфа,
БНИИСХ. 2003. 332с.
7.
Хайруллина Н.И. Черно-пестрая порода крупного рогатого скота на рубеже веков:
монография / Н.Г.Фенченко, Н.И.Хайруллина.-Уфа: БНИИСХ, 2000.-С.30-60.