Тимофійчук І.Р.
Буковинський державний медичний
університет
Активність
глюкозо-6-фосфатдегідрогенази за умов розвитку експериментального цукрового
діабету у щурів різних вікових груп.
Постійність
рівня глюкози в крові найважливіша умова підтримання нормальної життєдіяльності
організму. Нормоглікемія є результатом узгодженої роботи нервової системи,
гормонів і печінки. Інсулін прискорює надходження моноцукрів в інсулінзалежні
тканини, особливо глюкози. Інсулін активує використання глюкози клітинами
шляхом:
-
активування і індукції синтезу ключових ферментів гліколіза (глюкокінази,
фосфофруктокінази, піруваткінази);
-
збільшення включення глюкози в пентозофосфатний цикл (активування
дегідрогеназ глюкозо-6-фосфата, 6-фосфоглюконата);
-
підвищення синтезу глікогену за рахунок стимуляції утворення глюкозо-6-фосфата.
Глюкозо-6-фосфат дегідрогеназа (Г-6-Ф-ДГ; G6PD) — цитозольний фермент,
який входить в пентозофосфатний,
метаболічний шлях і забезпечує утворення клітинного НАДФ-H з НАДФ+. НАДФ-H і необхідний для підтримання відновленого глутатіона в клітинах, синтезу
жирних кислот і ізопреноїдів.
Інтенсивність
пентозофосфатного шляху залежить від функціонального стану тканин і від
гуморального статусу. Швидкість циклу регулюється концентрацією НАДФН.
Дегідрогенази пентозофосфатного циклу чутливі до змін величини співвідношення
НАДФ/НАДФН: при його величині 0,02 активність дегідрогенази в печінці
максимальна, а при величині 0,01 знижується на 90%. Інтенсивний
пентозофосфатный цикл відбувається в еритроцитах, що пов’язано з необхідністю
НАДФН-залежного відновленого глутатіона кофактора глутатіонредуктази
еритроцитів.
Пригнічення активності Г-6-Ф-ДГ в еритроцитах пояснюється
наявністю двох процесів, які приводять до втрати активності ензима. По-перше це
пошкодження молекул ферменту вільними радикалами, які підсилено виробляються в
умовах гіпоксії, а по друге глибокі порушення тіол-дисульфідної рівноваги в
клітинах і порушення активності Г-6-Ф-ДГ, як тіолзалежного ферменту. Тому
дефекти обумовлені зниженням активності Г-6-ФДГ, роблять НАДФH-залежний антиоксидантний захист клітин неефективним.
З
віком падає активність ферментів тканин мозку щурів - АТФаз, трансфераз. Найбільшу цікавість викликають
дані відносно активності різних ферментів під час старіння: в аорті знижуються
окисно-відновні процеси, в яких беруть участь: лактат-, малат-, гліцерол-3-фосфат,
глюкозо-6-фосфат-дегідрогеназа і флавінові ферменти - сукцинатдегідрогеназа, НАДН-дегідрогеназа.
Цукровий діабет, який приводить до гіперглікемії,
збільшення АФК і зниження активності ферменту Г-6ФДГ на фоні зростання рівня
глікозильованого гемоглобіна приводить до активації поліолового шляху обміну
глюкози і як наслідок до розвитку процесів нейродегенерації на фоні підвищення
рівня продуктів ПОЛ і зниження рівня Г-6-ФДГ.
На сьогоднішній день відомо, що одним із механізмів
пошкодження клітин при надмірному надходженні в клітину глюкози є активація
поліолового шляху обміну глюкози. Так при високих концентраціях глюкоза за допомогою
фермента альдозоредуктази перетворюється в сорбітол. Останній при участі
фермента сорбітолдегідрогенази метаболізується у фруктозу з утворенням NADH. Порушення вуглеводного обміну і гіперглікемія, яка
розвивається до клінічних проявів діабету, приводить до збільшення обміну
глюкози по поліоловому шляху і накопиченню сорбітола і осмотичному набряку
тканини, що стає причиною дегенеративних процесів в тканинах.
Старіння
є тим процесом, який модифікує нейрохімічний статус більшості тканин організму,
а цукровий діабет (ЦД) вважається процесом, який може бути аналогічним до
прискореного старіння - при цьому захворюванні зростає схильність до
дегенеративних станів.
Ми поставили за мету
визначити активність ферменту Г-6-ФДГ
у щурів різних вікових груп за умов розвитку експериментального цукрового
діабету.
Дослідження
проведено на нелінійних лабораторних щурах-самцях одно (молоді тварини),
п'ятимісячного (дорослі тварини) та вісімнадцятимісячного (старі тварини) віку.
Для відтворення цукрового діабету (ЦД) 2 типу за методом А.М. Ульянова і Ю.А.
Тарасова використовували протамін сульфат, який вводили внутрішньом'язево (в/м)
дозою 1 мм/кг 2 рази на день протягом 14 діб. Активність Г-6-ФДГ визначали спектрофотометрично
за збільшенням оптичної густини розчину, що обумовлено зростанням кількості
НАДФН2 у процесі ферментативної реакції.
В результаті
проведених експериментів виявлено (див. рис.), що у старих щурів по зрівнянню
із дорослими, і з молодими тваринами знижується активність ферменту Г-6-ФДГ, тоді як у середньої вікової
групи подібних змін не відмічалось, активність ферменту залишалась на рівні
молодих тварин. Подібні зміни активності ферменту Г-6-ФДГ узгоджується з даними літератури, і, можливо, є наслідком
зростання рівня активних форм кисню в старіючих тканинах. Експериментальний
цукровий діабет у щурів молодої і дорослої вікових груп викликав вірогідне
зростання вивчаємого показника в 1,4 рази і в 1,2 рази, що може бути наслідком
активації системи антиоксидантного захисту, а також відповідно до останніх
досліджень - посилення активності Г-6-ФДГ є одним із чинників захисту клітин
від наростаючого окисного стресу і може підтримувати рівень біоактивності
монооксиду нітрогену таким чином, щоб запобігти дисфункції. Літературні дані
вказують, що концентрація позаклітинної глюкози регулює експресію гена
індуцибельної NO- синтази (iNOS) і
продукування NO через Г-6-ФДГ-залежний
механізм. У щурів старої вікової групи відмічалось вірогідне зниження рівня
Г-6-ФДГ по зрівнянню із такими показниками у молодих і дорослих тварин із
цукровим діабетом в 1,2 та в 1,1 рази відповідно, та в 0,9 рази по зрівнянню з
показниками активності ферменту у щурів старшої вікової групи без цукрового
діабету, що по всій ймовірності є наслідком активації процесів перекисного
окиснення ліпідів на фоні зниженої активності ферментів антиоксидантного
захисту.
Рис.
Активність
Г-6-ФДГ в плазмі крові щурів за умов експериментального цукрового діабету у
щурів різних вікових груп
(M±m;
n=10)
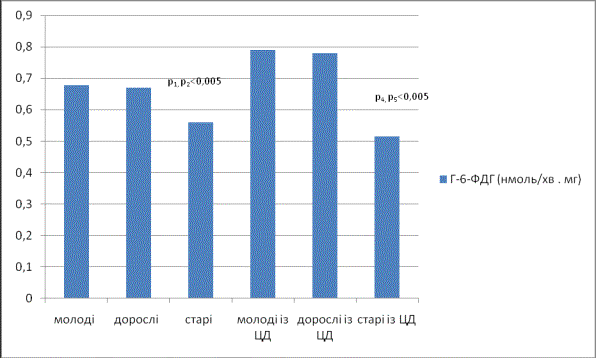
Примітка: р – статистично достовірна
різниця між дорослими та молодими тваринами; р1 – статистично
достовірна різниця між старими та молодими тваринами; р2 –
статистично достовірна різниця між старими та дорослими тваринами, р3 -
статистично достовірна різниця між молодими та дорослими тваринами із ЦД, р4
– статистично достовірна різниця між старими та молодими тваринами із ЦД; р5
– статистично достовірна різниця між старими та дорослими тваринами із ЦД.