д.м.н. Кокуркин
Г.В.
Чувашский государственный университет им. И.Н. Ульянова
г. Чебоксары
Математический анализ
энергетического состояния
клетки в организме
Каждая клетка
представляет собой миниатюрную саморегулирующуюся биоэнерго-информационную
систему и обладает всеми свойствами живого организма. Если представить клетку
как живую систему – она осуществляет обмен веществом и энергией, размножается и
передает по наследству свои признаки, реагирует на изменения внутренней и
внешней среды. Она является начальной ступенью организации, обладающей всеми
свойствами, наименьшей структурной и функциональной единицей живого.
Функциональная
система различного уровня организации строится по принципу саморегуляции.
Саморегуляция в функциональных системах приобретает специальную направленность.
Здесь отклонение результата деятельности функциональной системы от уровня,
обеспечивающего нормальный метаболизм (жизнедеятельность) организма (или
клетки) и его адаптацию к окружающей среде, является стимулом к мобилизации
необходимых элементов системы для обеспечения этого результата [5].
В каждой клетке
тела заложена генетическая информация, достаточная для того, чтобы был
воспроизведен весь организм. Эта информация записана в структуре дезоксирибонуклеиновой
кислоты (ДНК) и заключена в генах, расположенных в ядре.
Природа снабдила
клетку многими устройствами и механизмами. Границей клетки является
плазматическая мембрана. Она представляет собой двумерный жидкокристаллический
раствор глобулярных белков в липидах. Структурную основу мембраны составляют
липиды, среди которых преобладают фосфолипиды. Белки в мембранах различных
клеток выполняют определенные функции: они являются, например, ферментами или
транспортными белками. Кроме того, в состав мембран входят стероиды, гликопротеиды
и некоторые неорганические соли.
Основная
структура всех мембран представляет собой два параллельных слоя липидов
(биомолекулярный слой). Мембранные липиды – амфипатические молекулы – имеют гидрофобную
часть и гидрофильную часть. Такие молекулы образуют на водной поверхности
мономолекулярный слой. В водном окружении и в клетке образуются биомолекулярные
слои: гидрофобные части различных молекул повернуты дальше от водного
окружения, т.е. друг к другу, и удерживаются вместе сильными гидрофобными
взаимодействиями и слабыми силами Лондона-Ван-дер-Ваальса (2, 4). Вначале
казалось, что мембрана просто ограничивает и защищает внутреннее содержимое
клетки, пассивно обеспечивая поступление сюда необходимых веществ и выброс
отходов. Но энергетическая система всех клеток построена одинаково. Поэтому,
если бы мембраны клеток были просто ограничивающими оболочками, то, например,
сигнал к усилению деятельности клеток печени без препятствий передавался бы
всем клеткам тела, а это вызвало бы хаос. В действительности же оболочка каждой
клетки – мембрана – построена таким образом, что она воспринимает только нужные
ей сигналы.
Эти рецепторы
построены на восприятие одних сигналов и нечувствительны к другим. В
соответствии информации, поступившей с рецепторов мембраны, клетка меняет свою
активность, скорость процесса деления и т.д. Так, благодаря мембране клетка
отвечает только на нужную ей информацию.
Молекулы в
мембранах непрерывно меняются местами между собой и с молекулами, находящимися
по соседству в жидкостях. Мембрана как бы находится в динамическом равновесии с
жидкостной средой. Такое состояние делает мембраны проницаемыми для молекул
различных веществ, которые нужны клетке для жизненных процессов.
Внутри клетки
идут процессы химических преобразований, перемещения и непрерывной замены
молекул в клеточных структурах. Эти процессы обычно называются обменом веществ.
Обмен осуществляется во всех клеточных структурах, он обеспечивает их сохранение
и стабильность.
Живое состояние
не структура, а процесс. Структуры живого не стабильны, а постоянно разрушаются
и строятся заново. Это обновление протекает с различными скоростями.
Каждая свободно
идущая химическая реакция
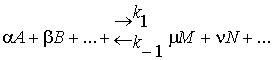
(например, ![]() ) достигает, в конце концов, состояния равновесия (термодинамическое
равновесие), когда скорость прямой реакции
) достигает, в конце концов, состояния равновесия (термодинамическое
равновесие), когда скорость прямой реакции ![]() и скорость обратной
реакции
и скорость обратной
реакции ![]() равны друг другу.
равны друг другу.
Согласно закону
действующих масс
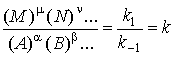 .
.
Этот закон
описывает состояние равновесия в замкнутой системе. Константа равновесия k
зависит от температуры. Катализатор (фермент), напротив, не оказывает влияния
на k, т.е. на положение термодинамического равновесия; он влияет только на
время, необходимое для достижения равновесия.
Если, однако, на
одной стороне в реакцию постоянно вступают новые количества исходных веществ, а
на другой стороне удаляются продукты реакции, то мы имеем открытую химическую
систему. В этом случае может быть достигнуто динамическое равновесие, когда при
постоянном потоке веществ, участвующих в реакции, устанавливаются их стационарные
концентрации. Это состояние называется стационарным состоянием. Все клетки
представляют собой открытые химические системы, так как они постоянно поглощают
новые исходные вещества и выделяют продукты обмена [2, 4].
Молекулы
веществ, необходимых для самой клетки или нужных другим клеткам организма,
синтезируются в эндоплазматической сети и в аппарате Гольджи. Молекулы веществ,
производимых для нужд данной клетки, непосредственно поступают в цитоплазму, а
затем употребляются в клеточных структурах для синтеза и сборки биополимеров.
Молекулы
веществ, использованные в клетке и предназначенные «на экспорт», упаковываются
в аппарате Гольджи в компактные гранулы и в таком виде через эндоплазматическую
сеть выделяются в межклеточную среду.
Энергия,
необходимая для процессов синтеза, сборки и для осуществления транспорта,
вырабатывается в митохондриях.
Энергия,
освобождающаяся при распаде органических веществ, не используется в клетках
сразу для осуществления работы, а сначала запасается в форме высокоэнергетических
промежуточных соединений – как правило, в форме аденозинтрифосфата (АТФ).
Фосфатные группы
соединены между собой так называемыми «высокоэнергетическими» связями.
В результате
гидролитического отщепления концевой фосфатной группы из АТФ образуется
аденозиндифосфат (АДФ). При этом освобождается около 30 кДж/моль энергии:
![]() ;
; ![]() кДж/моль.
кДж/моль.
Наоборот, при синтезе АТФ из
АДФ и H3PO4 должно быть затрачено около
30 кДж/моль.
При отщеплении
второй фосфатной группы получается аденозинмонофосфат (АМФ):
![]() ;
; ![]() кДж/моль.
кДж/моль.
Гидролитическое отщепление
последней фосфатной группы освобождает лишь –13 кДж/моль.
Таким образом, в
биохимическом преобразовании энергии можно выделить два основных этапа: 1)
синтез пирофосфатных связей АТФ и 2) использование их для совершения работы.
АТФ чрезвычайно быстро обновляется. У человека каждая молекула АТФ расщепляется
и вновь регенерируется 2400 раз в сутки, так что средняя продолжительность ее
жизни менее 1 мин [2, 4].
Человеческий
организм существует в состоянии неравновесного порядка, когда энергетические
потери компенсируются за счет энергии топлива (пищи) и окислителя (воздуха).
Клетка
представляет собой открытую биоэнерго-информационную систему, т.е. обменивается
веществом и энергией с окружающей средой. Различают внешний обмен – поглощение
и выделение веществ, и внутренний обмен – химические превращения этих веществ в
клетке. Обмен веществ (метаболизм) включает непрерывный синтез (анаболизм) и
частичное разрушение (катаболизм) компонентов живого организма.
Любая работа,
выполняемая клеткой – поглощение веществ («осмотическая» работа), двигательные
реакции, расширенное воспроизведение живой материи, – требует дополнительной
энергии (функциональная энергия). Всю эту энергию доставляют высокоэнергетические
соединения, прежде всего АТФ.
Высокоэнергетические
соединения, такие как АТФ, получают свой запас энергии благодаря диссимиляции,
т.е. экзергоническим процессом расщепления органических веществ (углеводов,
жиров, белков) – процессам катаболизма. Органические вещества, необходимые для
построения живой материи, образуются в ходе ассимиляции.
Такие изменения
энергетического состояния клетки в сутки можно выразить математическим
выражением:
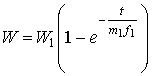 , (1)
, (1)
где 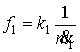 – функция нервной
системы на выработку в клетке АТФ, k1 – постоянная,
– функция нервной
системы на выработку в клетке АТФ, k1 – постоянная, ![]() – скорость выработки
АТФ (нг/с), m1 – количество АТФ в клетке, t1 – характерное время,
необходимое для перехода клетки из одного энергетического состояния W0 в другое W1, которое получается
логарифмированием выражения (1):
– скорость выработки
АТФ (нг/с), m1 – количество АТФ в клетке, t1 – характерное время,
необходимое для перехода клетки из одного энергетического состояния W0 в другое W1, которое получается
логарифмированием выражения (1): ![]() .
.
В следующий
момент вследствие затрат имеет место убыль энергии, вызванная расходом АТФ,
тогда это можно выразить следующим выражением:
![]() , (2)
, (2)
где 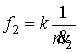 – функция нервной
системы на расход АТФ, k2 – постоянная,
– функция нервной
системы на расход АТФ, k2 – постоянная, ![]() – скорость расхода
АТФ в клетке (нг/с), m1 – количество АТФ в клетке, t2 – характерное время, необходимое
для перехода клетки из одного энергетического состояния W1 в другое W2, равное
– скорость расхода
АТФ в клетке (нг/с), m1 – количество АТФ в клетке, t2 – характерное время, необходимое
для перехода клетки из одного энергетического состояния W1 в другое W2, равное ![]() .
.
При достижении
определенной концентрации АТФ в клетке скорость его выработки уменьшается
(покой).
![]() – характерное время
достижения энергетического уровня W2 в клетке, ниже которого
поступления энергетических веществ в организм происходит реакция нервной
системы на пополнение (ускорение процессов выработки) АТФ.
– характерное время
достижения энергетического уровня W2 в клетке, ниже которого
поступления энергетических веществ в организм происходит реакция нервной
системы на пополнение (ускорение процессов выработки) АТФ.
Далее,
выработкой АТФ энергетическое состояние клетки вновь увеличивается за время ![]() , переходом клетки из состояния W2 в W1.
, переходом клетки из состояния W2 в W1.
Вследствие
затрат клетка в дальнейшем вновь переходит из состояния W1 в состояние W2 за время ![]() , аналогичное времени t2.
, аналогичное времени t2.
При переходе
клетки из активного состояния в пассивное (сна) энергетическое состояние клетки
описывается W0 и необходимое для этого время ![]() .
.
Во время
пассивного состояния t6–0 энергетическое состояние
клетки почти не меняется и сохраняется на уровне W0.
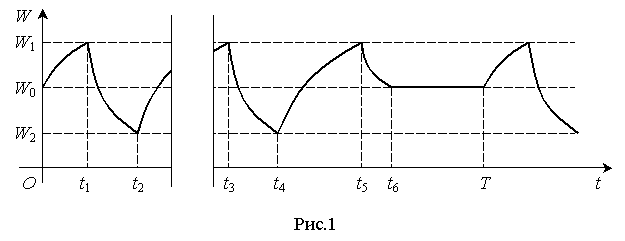
Эти процессы
можно представить графически в координатах W(t) (рис.1).
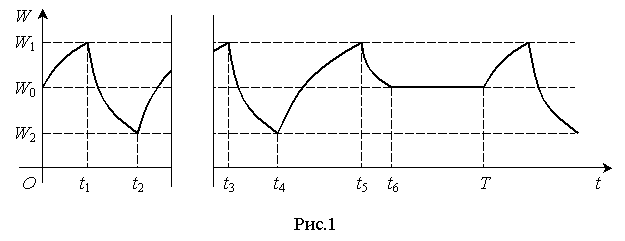
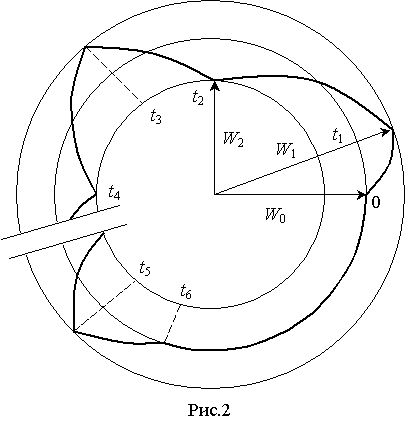
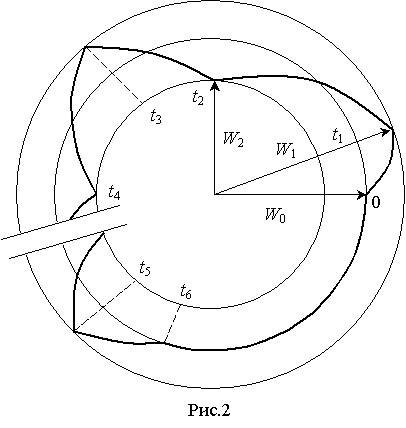
Релаксационные
процессы можно представить в координатах W(t) на фазовой плоскости, где ![]() – предел изменения
энергетического состояния нормально функционирующей в организме клетки в
течении суток (рис.2). Аналогичные результаты получены в экспериментальных
исследованиях и другими авторами (1), однако аналитические зависимости не описаны.
– предел изменения
энергетического состояния нормально функционирующей в организме клетки в
течении суток (рис.2). Аналогичные результаты получены в экспериментальных
исследованиях и другими авторами (1), однако аналитические зависимости не описаны.
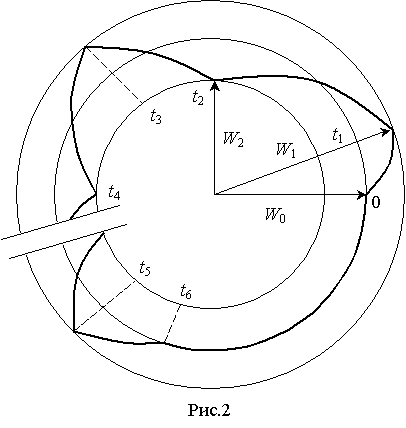
Таким образом, в
клетке происходят следующие функции, которые составляют обмен веществ:
транспорт из внеклеточного пространства исходных веществ для жизнедеятельности
клетки, получение энергии из этих веществ, синтез и сборка биополимеров по генетической
программе, разложение ненужных веществ и выведение этих конечных продуктов за
пределы клетки.
Литература:
1. Комаров, Ф.И.
Хронобиология и хрономедицина / Ф.И. Комаров, С.И. Рапопорт. - Москва.: Триада,
2000. - 488 с.
2. Мамонтов,
С.Г. Биология / С.Г. Мамонтов // Учебное пособие. - М.: Дрофа, 1999. - 480 с.
3. Либберт, Э.
Основы общей биологии / Под ред. Э. Либберта. - Москва, 1982. - 440 с.
4. Судаков, К.В.
Нормальная физиология / Под ред. К.В. Судакова. – М.: Медицинское информационное
агентство, 1999. - 718 с.
5. Судаков, К.В.
Функциональные системы организма / Под ред. К.В. Судакова. –М.: Медицина, 1987.
- 432 с.