Рост и развитие плода на разных стадиях онтогенеза
Н.Г.Фенченко, Н.И.Хайруллина, Р.Ф.Галимов,
А.З.Шайхутдинова, Ф.М.Гафарова, Н.В.Ильина - аспирантка
Известно, что растущий и развивающийся организм
животного представляет собой сложную систему, основу существования которого
определяет обмен веществ с окружающей средой. Взаимосвязанные
физико-химические, биохимические, структурные и функциональные изменения,
протекающие в нем, обеспечивают его неразрывное самообновление (Н.Г.Фенченко,
2007). На разных стадиях эмбриогенеза животных наблюдается довольно краткий
период резкого повышения метаболизма, а также интенсивности и полноценности
самообновления, что в некоторой степени является показателем скорости роста
плода в зависимости от пола (рис. 1).
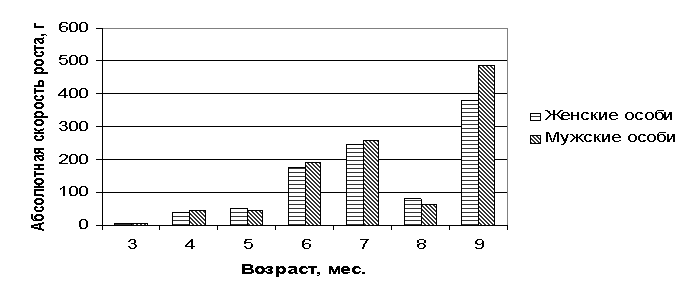
Рис. 1 Динамика абсолютной скорости роста плода
С целью изучения биологических закономерностей
весового роста в опытах на эмбрионах черно-пестрой породы в зависимости от пола
и возраста по формуле С.Броди вычисляли абсолютную и относительную скорость
роста. Изучены экстерьерно-конституциональные особенности плода в разные
возрастные периоды внутриутробного развития с последующим вычислением
соответствующих индексов.
Превосходство мужских особей над женскими по
интенсивности роста довольно четко прослеживалось вплоть до шестого месяца
развития плода 0,25…14,75 г. Однако на пятом и восьмом месяце фазы роста и
развития онтогенеза превосходили уже женские особи (5,5…15,5), тогда как во все
последующие явное преимущество интенсивности развития плода было на стороне
мужских особей (6,5…88,21). Причем
самые высокие показатели абсолютной скорости независимо от пола были на седьмом
(246…258 г), девятом (379,3…467,5 г) месяце эмбрионального периода их развития.
Наиболее высокая изменчивость абсолютной скорости роста была у самцов и самок
на третьем (28,6...31,5%) и на восьмом (26,9...58,2%), тогда как у самцов
стабильно высокие показатели наблюдались еще и в четырех, шести семимесячном возрасте (15 5...
16,9%). На скорость абсолютного роста, как
отмечает А.А. Малигонов (1968), во многом оказывает влияние порода.
Данное явление объясняется тем, что в процессе
развития организма имеет место ритмичность показателей скорости и роста. Если
в этот период качественные изменения сопровождаются расчленением целого на
части, то такие изменения в живой материи характеризуют другую сторону его
развития.
Таким образом, абсолютный прирост растущих
животных был не значителен, затем увеличения достигли максимума, после чего
уменьшались, а максимум относительного прироста приходился на более ранние
ступени их развития (рис. 2).
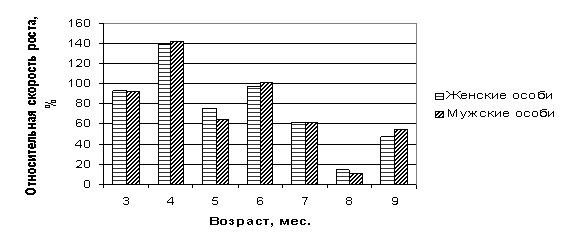
Рис. 2 Динамика относительной скорости роста
Относительная скорость роста, вычисленная по С.
Броди, (1945) показывает, что эмбрионы развивались независимо от пола с
одинаковой закономерностью строго с учетом ритма, где их интенсивность
чередовалась с максимальным приростом, преимущество было на ранних стадиях
эмбриогенеза. Причем наиболее высокие показатели относительной скорости роста
прослеживались на четвертом (138,7...141,8%) и шестом месяцах эмбрионального
периода (97,5... 100,7%). Наиболее низкие показатели скорости роста приходится
на восьмой месяц онтогенеза крупного рогатого скота (11,35... 14,69%), тогда
как в остальные месяцы она была довольно высокой (47,7...92,8%).
Таким образом, скорость роста плода во многом
обуславливается преимущественно в формировании и развитии отдельных его
органов и тканей.
Конституция и экстерьер,
как и другие фенотипические признаки, формируются под влиянием генотипа и
условий жизни организма. Совместные действия этих факторов осуществляется в
процессе индивидуального развития животных. Внутренние органы и ткани
организма, создавая известный баланс в обмене веществ, в конечном итоге
определяют корреляционные связи между разными особенностями экстерьера и
физиологическими свойствами организма (Н.Г. Фенченко, Н.И. Хайруллина 1995).
Между тем промеры,
характеризующие развитие плода и его частей в длину, изменялись по несколько
иной закономерности, несмотря на сохраняющуюся их ритмичность. Более резкое
увеличение начиналось у мужских особей с прямой, а у женских - косой длины
туловища (11,3...11,9 см) (10,9...14,5 см), семимесячных соответственно
(14,5...8,3 см) и (15,8...7,0 см) девяти мес. (7,8...11,5 см) и (7,3...10,5
см). Особо следует выделить изменения промеров характеризующих развитие
грудной клетки у мужских плодов, где у четырехмесячных глубина, ширина груди
увеличилась на 12,0...3,6 см., а обхвату пятимесячных на 3,7; 1,34; 19,3 см.
По женским особям при
общей закономерности имели место некоторые свои особенности. Так на четвертом
месяце разница в увеличении данных промеров составила 11,3...3,7 см и 12,25 см,
пятимесячных соответственно значительно меньше, чем предыдущие мужским
(2,0...5,0 и 5,7 см), у семимесячных происходил некоторый рост промеров, чем
предыдущих на 3,0; 2,6 и 9,0 см а восьмимесячных соответственно 0,75; 1,25 см и
существенное увеличение наблюдалось по обхвату груди (20,0 см), последний
ровно на половину снизил темп своего роста 10,2 см.
Тем не менее, процесс
формирования грудной клетки связан с ростом и развитием данных промеров по
сравнению с остальными Формирование задней трети туловища у плодов изучаемой
нами молочной породы также представляет особый интерес, так как их развитие
существенным образом отличается от представителей мясного направления
продуктивности.
По ширине в маклоках,
седалищных буграх и полуобхвату зада мужские особи в четыре месяца своего
развития существенно превосходили трехмесячных на 5,65; 3,92 и 6,5 см,
семимесячных соответственно на 4,25; 150 и 6,25 см и девятимесячных на 2,5; 1,0
и 9,5 см.
По женским особям данные
различия были как в возрастном, так и количественном измерении по сравнению с
мужскими.
В четыре месяца
увеличение составило 4,07;. 4,37 и 6,5 см, в пять соответственно 5,8; 0,88 и
4,0 см, затем семимесячных 1,35; 0,66 и 6,5 см и девятимесячных 1,75; 1,0 и
9,25 см. Тем не менее, довольно существенно изменялись промеры по ширине в
седалищных буграх и обхвату бедра, где у мужских на четвертом месяце разница
составила 10,75 и 12,25 см, пятимесячных соответственно 7,0 и 8,25 см,
семимесячных 9,25 и 4,0 см и девятимесячных 9,0 и 6,25 см, а женских
соответственно 10,75... 13,45 см; 6,85...6,45 см; 10,25...3,75 см и 8,75...6,25
см (рис. 3).
Таким образом, анализ
приведенных данных свидетельствует о том, что показатели линейного роста у
животных изменялись и имели свои закономерности в эмбриональный период,
обуславливающие формирование органов и тканей с учетом их периодичности.
Рис.3. Взятие промеров у
плода пятимесячного возраста
Поэтому
выявление особенностей формирования органов и тканей животных путем вычисления
соответствующих индексов, наряду с изменением его массы является одним из важных
биологических приемов при совершенствовании породы. Так же как и промеры,
индексы дают возможность выявить те моменты, когда организм животного
нуждается в большей степени воздействия на
него через организм матери. Анализ полученных данных показывает, что индексы
телосложения существенно изменялись в процессе всех изучаемых фаз
эмбрионального периода, как у мужских, так
и у женских особей. Причем
почти все они изменялись крайне неравномерно, а поэтому установить какую
либо четкую закономерность на первом этапе не представляется возможным (рис.
4, 5).
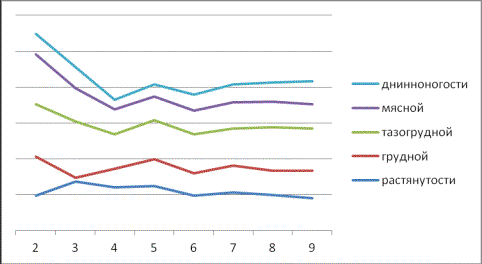
Рис. 4. Динамика индексов мужских эмбрионов в
зависимости от фазы развития
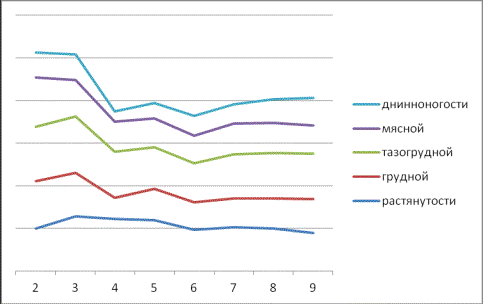
Рис. 5. Динамика
индексов женских эмбрионов в зависимости от фазы развития
В частности, на третье
месяце эмбрионального периода только три индекса – это растянутости,
перерослости и массивности существенно увеличились по сравнению с
двухмесячными, как у женских (24,16-37,60), так и у мужских (19,92-42,47), при CV=5,28-11,25
%.
Однако довольно сильное
изменение в сторону уменьшения имело место по мясному индексу (Грегори) у
женских на 3,25, а у мужских еще больше – на 45,23 при CV=11,29-12,22 %.
Снижение, но менее
значительное, имело место по грудному индексу (8,92-2,52) независимо от пола,
тогда как по тазогрудному наблюдалось незначительное увеличение (5,48-8,48).
Особо следует выделить
развитие таких показателей, которые довольно существенно снижались независимо
от пола, тогда как грудной – на 24,23 у женских и на 22,62 – у мужских особей,
широкозадости у женских на 13,12 и длинноногости независимо от пола на шестом
месяце (10,65-11,60).
По другим индексам имело
место их незначительное увеличение длинноногости соответственно (6,00-11,76) и
(4,6-8,07).
Изменчивость остальных
индексов была невысокой и почти одинаковой по фазам эмбрионального периода.
Такие резкие перепады в изменении индексов объясняются нарушением и
восстановлением относительного равновесия между процессами ассимиляции и
диссимиляции, наблюдающихся в онтогенезе животных. При этом неравномерность
онтогенеза во многом связана, как уже отмечалось ранее, уменьшением с возрастом
развития отдельных органов и тканей плода.
Между
тем, по мнению Н.Руденко и др. (1981), объективным показателем эмбрионального
развития теленка является его живая масса при рождении. На продолжительность
внутриутробного развития приплода существенное влияние оказывает возраст и
живая масса матерей при первой случке. Установлено, что наибольшую живую массу
при рождении имели телята, полученные от матерей, первый раз отелившихся в
возрасте 27 месяцев. Живая масса новорожденных телят прямо пропорционально
связана с продолжительностью эмбрионального периода. Так живая масса приплода,
полученного от матерей в возрасте 27 месяцев составила 30,7…35,3 кг при
продолжительности периода выращивания 282,7…285,3 дней. Период стельности у
нетелей, слученных в 12 и 15 месяцев, оказался по сравнению с контрольными
короче на 3,7…5 и 1,5…3 дня. Этим объясняется и меньшая живая масса телят при
рождении 26,3…29,0 и 28,7…32,7 кг против 30,7…35,3 кг. Причем большинство
первотелок, отелившихся в возрасте 21 месяцев, довольно тяжело переносили роды
и в семи случаях из них имели место травматические повреждения слизистой
влагалища с кровотечениями. Здесь же наблюдалось задержание последа до 19,5 часов,
и при тяжелых родах проводилось родовспоможение.
Таким образом, с учетом особенностей
эмбрионального развития представляется возможность активно влиять на разработку
инновационных технологий, включающих рациональное кормление, содержание
стельных коров, при этом активно и целенаправленно вмешиваться в процессы
формирования продуктивных качеств выращиваемого и откормочного молодняка,
являющимся одним из резервов увеличения количества и улучшения качества
получаемой от них продукции.