Чернетченко Д.В., Моцний М.П., Боцьва Н.П., Єліна
О.В.
Дніпропетровський
національний університет ім. Олеся Гончара, Україна
МАТЕМАТИЧНА МОДЕЛЬ РОСЛИННОЇ КЛІТИНИ ІЗ ЗДАТНІСТЮ ГЕНЕРУВАТИ АКТИВНІ ЕЛЕКТРИЧНІ ПОТЕНЦІАЛИ
У наш час залишається актуальною
задача контролю розвитку рослин в штучних умовах їх вирощування. Зміна
функціонального стану рослин під дією зовнішніх змінних факторів може
сприятливо впливати на їх розвиток. На жаль, переважна більшість існуючих
засобів контролю життєдіяльності так чи інакше впливає на стан рослин у процесі
проведення вимірювань.
Запропоновано інший підхід до
вирішення цієї задачі – модельні дослідження біоелектричних показників стану
рослин на еквівалентних математичних моделях, що побудовані на отриманих у
реальних умовах життєдіяльності рослин експериментальних даних. Встановлено, що
потенціали дії клітин, викликані подразниками різної природи – температурними,
світловими та електричними стимулами, – значно відрізняються як за формою, так
і за параметрами, тому об’єктивно
характеризують поточний стан рослинних клітин
[1,2].
Для аналізу можливих
внутрішньоклітинних механізмів виникнення та динаміки зареєстрованих біоелектричних
потенціалів розроблено математичну модель клітини із здатністю генерувати
активні електричні потенціали у відповідь на світлові імпульси.
Результати
експериментальних досліджень на рослинах свідчать, що короткий світловий
імпульс приводить до виникнення значного потоку йонів Н+,
який пояснюється збільшенням швидкості пасивного потоку Н+ до
цитоплазми, гальмуванням активного транспорту СО2 із
середовища в темнових реакціях фотосинтезу, а також можливістю поглинання
світла хлоропластами із наступною активацією іонних каналів Н+
та подальшою зміною pH на поверхні клітини [3]. Для опису динаміки іонних
струму гідрогену ![]() та трансмембанного потенціалу
та трансмембанного потенціалу
![]() під час світлової
стимуляції запропоновано підхід на основі транс-мембранного транспорту
під час світлової
стимуляції запропоновано підхід на основі транс-мембранного транспорту
![]() ,
,
де ![]() ,
, ![]() – потенціал рівноваги та електрична провідність іонних
каналів H+.
– потенціал рівноваги та електрична провідність іонних
каналів H+.
Зміна загального мембранного
потенціалу описана рівнянням моделі Ходжкіна-Хакслі [4]
![]() ,
, ![]() ,
,
де ![]() – густина s-го йонного струму через мембрану; С – питома електроємність
мембрани.
– густина s-го йонного струму через мембрану; С – питома електроємність
мембрани.
Системи первинного (протонна й
кальцієва АТФази) і вторинного (![]() – сімпортер і
– сімпортер і ![]() – антипортер) транспорту описані як системи, що можуть
перебувати у двох станах: вільному та зв’язаному з іоном. Потік через ці
транспортні системи знайдено як стаціонарний розв'язок системи рівнянь
– антипортер) транспорту описані як системи, що можуть
перебувати у двох станах: вільному та зв’язаному з іоном. Потік через ці
транспортні системи знайдено як стаціонарний розв'язок системи рівнянь
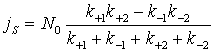 ,
,
де ![]() – загальна концентрація ферменту; k+1, k–1
– константи
прямого (потік до клітини) та
зворотного (потік у зовнішнє середовище) переходу "вільний стан – стан, зв’язаний з іоном";
k+2, k–2
– константи
переходу "стан, зв’язаний з іоном – вільний стан".
– загальна концентрація ферменту; k+1, k–1
– константи
прямого (потік до клітини) та
зворотного (потік у зовнішнє середовище) переходу "вільний стан – стан, зв’язаний з іоном";
k+2, k–2
– константи
переходу "стан, зв’язаний з іоном – вільний стан".
Константи для протонної помпи
описані рівняннями
![]() ,
,
![]() ,
,
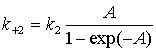 ,
,
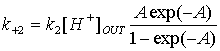 ,
,
де k1, k2 – константи, не залежні від мембранного
потенціалу й концентрації протонів; [H+]IN, [H+]OUT
–
концентрації протонів у цитоплазмі й апопласті; ![]() – енергія, що
вивільняється при гідролізі АТФ; А = (FEM/RT).
– енергія, що
вивільняється при гідролізі АТФ; А = (FEM/RT).
Для опису функції активації клітинних механізмів F(E) застосовано лінійну апроксимацію
на основі експериментальних даних [1,2]
|
|
|
0, |
|
|
F(E)= |
|
|
|
|
|
1, |
|
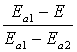
де ![]() – потенціали активації та
інактивації йонних каналів.
– потенціали активації та
інактивації йонних каналів.
Рівняння для густини струму має
вигляд
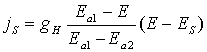 .
.
За зміною величин амплітуд
відповідей на основі отриманих експериментальних даних обчислено значення
провідності мембрани для іонів гідрогену (протонний транспорт), яке відповідає
значенню провідності протонних каналів мембрани [5]. Це достатньо достовірно вказує
на походження електричних відповідей рослинних клітин, зареєстрованих в
експериментах із світловою стимуляцією. Такі відповіді є потенціалами дії, в
основі яких лежить транспорт іонів H+ [5,6]. Незначне відхилення
розрахованих під час моделювання електричних провідностей від табличних значень
для відповідних іонів обумовлено наявністю у клітині струмів інших типів, а
також пасивного іонного струму витоку.
Таким чином, у результаті
визначення на базі отриманих експериментальних даних параметрів математичної
моделі клітини із здатністю генерувати активні електричні потенціали
встановлено, що в основі електричної відповіді рослинної клітини на фотостимули
лежить селективна зміна електричної провідності клітинної мембрани для іонів Н+,
що доводить навність тісного зв’язку між електричними потенціалами на поверхні
рослини та внутрішньоклітинними процесами.
Література
1. Вплив фотостимуляції на
біоелектричну реакцію листя кукурудзи / М.П.Моцний та ін. // Вісник ДНУ.
Біологія. Екологія. 2011. Т.2. – С.103-108.
2. Автоматизована система реєстрації біоелектричних
потенціалів / Д.В.Чернетченко,
М.П.Моцный та ін. // Вісник ДНУ. Біологія. Екологія. 2013. Т.2. – С.70-75.
3. Davies E. New functions for electrical signals in plants // New Phytol. 2004.
V. 161. – P. 607-610.
4. Hodgkin
A.L., Huxley A.F. A quantitative description of membrane current and its
application to conduction and excitation in nerve // J.
Physiol. 1952. V. 117. – Р. 500-544.
5. Биофизика / В.Ф.Антонов и др.– М.: Владос, 2000. – 288 с.
6. Распространение
вариабельного потенциала; индуцированного ожогом семядольного листа проростка
тыквы / В.А.Воденеев, Р.В.Мамонов и др. // Вестник ННГУ.
2007. № 2. – С. 122-126.