Екологія/3. Радіаційна безпека
соціально - екологічні
проблеми
Д.б.н. Чорна В.І.
Дніпропетровський державний аграрний
університет, Україна
Біологічна дія малих доз іонізуючої радіації на клітини ЦНС
Дослідження механізмів
залежності змін метаболізму від дози опромінення є актуальною проблемою
радіаційної екології як в загальнотеоретичному аспекті, так і можливості їх
практичного застосування за специфічних обставин, що виникли внаслідок аварії
на Чорнобильській АЄС [1]. Недостатньо вивчені і механізми розвитку радіаційних
зрушень і змін в організмі, які виконують важливу роль у патогенезі і перебігу променевих
уражень в забезпеченні компенсаторних та репараційних процесів. Актуальність
досліджень в цьому напрямку зросла після аварії на ЧАЄС, оскільки в наслідок
цієї катастрофи значні території стали зоною підвищеного екологічного та
біологічного ризику [2]. Прогнозування можливих біологічних ефектів, зумовлених
дією малих доз радіації на живі організми, викликає необхідність пошуку тестів,
яки б дозволили адекватно оцінити ступінь цього впливу. Особливого значення
набуває виявлення метаболічно важливих показників, визначення яких дозволить говорити
про зміни всієї системи клітинного метаболізму в умовах техногенного
радіоактивного забруднення. Одним із таких перспективних напрямків є
дослідження внутрішньоклітинних лізосомних цистеїнових протеаз та їх
інгібіторів. Як відомо, протеоліз є важливою ланкою в регуляції багатьох
біохімічних процесів [3]. Аварія на Чорнобильській АЄС виявила, що найменш дослідженими
при дії малих доз радіації є зміни метаболізму ЦНС. Згідно з концепцією первинної
активуючої дії малих доз [2] для ініціації адаптивної відповіді достатньо дуже
малої (1сГр) дози, яка порушує проникність мембран. Припускається наявність
існування нижнього порогу малих доз, які ініціюють адаптивну відповідь [1,2]. В
літературі обговорюється декілька можливих механізмів формування відповідної
реакції клітин на опромінення в малих дозах [2]. Незважаючи на принципові
відмінності, властиві цим підходам, їх об’єднує головне – прагнення пояснити
триггерний характер наявних змін. Актуальність вивчення лізосомних цистеїнових
катепсинів нервової системи визначається з одного боку тими функціями, які вони
виконують в клітині, а з другого – тією роллю, яку ці ферменти відіграють в
системі відповіді організму на дію шкідливих факторів оточуючого середовища.
З відкриттям явища гормезису
інтенсифікувались дослідження впливу низьких доз радіації [4]; а також з’ясування
відмінностей на молекулярному, клітинному і організменному рівнях у механізмах
дії радіації на біологічні об’єкти за малих та великих доз і потужностей.
Неможливість екстраполяції ефектів великих доз є очевидною і витікає з
характеру загальновідомих кривих виживання [5]. Загальною для всіх ефектів
малих доз радіації є активація різних ферментативних реакцій.
При дослідженні деяких ефектів в
діапазоні малих доз встановили залежність і від потужності дози, при чому форма
залежності може змінюватись разом з величиною дози, що свідчить про складність
самих явищ.
Метою даної роботи було
дослідження дозової залежності активності лізосомного цистеїнового катепсину L
в діапазоні малих доз для з’ясування адаптивної відповіді по зміні вільної
активності катепсину L у корі великих півкуль головного мозку і мозочку щурів.
За даними, наведеними у роботі [6], діапазон потужності дози, в якому проявляється
адаптивна відповідь у лімфоцитів людини, знаходиться від 1 до 25 сГр/хв.. При
меншій потужності дози активації не виникає із-за швидкої репарації ушкоджених
мембран, а при більшій – із-за превалюючого інгібіторного ефекту. Тому в своїх
дослідженнях ми використовуємо потужність дози опромінення 0,26 сГр/хв..
Умови експериментального радіаційного
опромінення наступні. Щурів опромінювали на апараті РУМ-17. Опромінювання
відбувалося за таких технічних умов: напруга 150 кВ, сила струму 6 мА, фільтри
0,5 мм Cu +2мм Cu, потужність дози 0,26 сГр/хв,
фокусна відстань1,86м. Дозові залежності визначали через 12 годин після
опромінення щурів за доз 5сГр, 10сГр, 20сГр, 25сГр. Ґрунтуючись на концепції наявності біохімічної та морфофункціональної
топографії головного мозку, а також їх взаємозв’язку у ЦНС [7] головний мозок розділяли на кору великих
півкуль і мозочок. Ці мозкові структури виділяються анатомічно, мають
морфологічні та функціональні особливості при здійсненні
інтегративно-синтетичної діяльності головного мозку. Використовували 10% екстрактів
тканин на 0,025М трис-буфері з рН 7,4, який містив 0,15М
Na Cl та 1М ЕДТО. Активність катепсину L визначали
по відношенню до азоказеїну, денатурованого 3м сечовиною. Статистичну обробку
результатів проводили загальноприйнятими методами.
За результатами досліджень при
опроміненні щурів в інтервалі доз від 0,05 Гр до 0,25 Гр з потужністю дози 0,26сГр/хв
визначаються відповідні зміни вільної активності катепсину L у структурах
головного мозку. Експериментальні дані по зміні вільної активності катепсину L
у корі великих півкуль головного мозку і мозочку в залежності від дози
опромінення представлені на рис. 1.
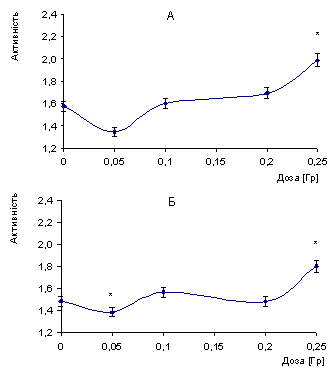 Рис.1.Залежність вільної
активності катепсину L у корі великих півкуль (А), мозочку (Б) головного мозку
щурів від дози опромінення (активність - в од. абсорбції при 366 нм/мг білка,
М±m, n= 6¸8). *- р < 0,05
порівняно з контролем.
Рис.1.Залежність вільної
активності катепсину L у корі великих півкуль (А), мозочку (Б) головного мозку
щурів від дози опромінення (активність - в од. абсорбції при 366 нм/мг білка,
М±m, n= 6¸8). *- р < 0,05
порівняно з контролем.
Підвищення активності ферменту
спостерігається після дози 0,1 Гр і за дози 0,25 Гр. Збільшення вільної
активності катепсину L у корі великих півкуль і мозочку набуває вірогідних
значень, і більше від контрольного рівня на 26% і 21,5% відповідно. З наведених
даних видно, що переломною у ефекті впливу на рівень вільної активності
катепсину L мозку є доза 0,1 Гр. Ейдус Л.Х (2) також зазначає, що для ініціації
адаптивної відповіді достатньо декількох секунд дії дози 1сГр. Радіаційні
ефекти таких впливів можуть викликати порушення проникності клітинних, а в
нашому випадку – лізосомальних мембран. Згідно з концепцією первинної
активуючої дії малих доз Ейдуса Л.Х., для ініціації адаптивної відповіді
достатньо дуже малої (1 сГр) дози, яка порушує проникність плазматичних
мембран, що спричиняє вихід із клітин і зниження внутрішньоклітинного пулу
різних низькомолекулярних субстратів.
В результаті ефекту неспецифічної
регуляції активності ферментів підвищується активність попередніх репаративних
ферментів і інтенсифікуються процеси їх відновлення. Порушення проникності,
викликані такою слабкою дією, швидко репаруються, і тим швидче, чим вони
слабкіші. Якщо швидкість репарації перебільшує швидкість виходу субстратів із
клітин, адаптивна відповідь не спостерігається. Це зумовлює існування нижнього
порогу малих доз, які ініціюють адаптивну відповідь. Наявність такого порогу
показана при аналізі адаптивної відповіді в клітинах китайського хом’яка із застосуванням
мікроядерного тесту.
З ростом дози зверх цього порогу
пошкодження мембранних функцій підвищується, репарація не встигає їх
компенсувати і в клітині виникає стан підвищеної активності, який спричиняє
адаптивну відповідь [2].
Відповідно до концепції Ейдуса
Л.Х. зміни вільної активності катепсину L, які спостерігаються у корі великих
півкуль і мозочку, мають поріг за дози 0,1 Гр, а стан підвищеної активності
клітин мозку припадає на дозу 0,25 Гр.
В експерименті різних авторів на
різних об’єктах показано, що суміжні з малими дози індукують процеси, які
спричиняють активацію геному і супроводжуються появою нових генних продуктів, у
тому числі білків, які в інтактних клітинах знайдені не були, а також відбувається
підвищення кількості ферментів, які репарують пошкодження ДНК. В основі
відповідної реакції клітини на опромінення в діапазоні від декількох мГр до 10
сГр лежить ініціація синтезу ферментів репарації, не зумовленої функціонуванням
SOS-системи. Більш того, на відмову від SOS-репарації адаптивна відповідь
приводить до зниження виходу мутацій [6].
Таким чином, нами встановлено, що
дозова залежність вільної активності катепсину L структур головного мозку має
нелінійний характер і може бути механізмом переключення клітин у новий режим
функціонування за дії малих доз радіації.
Література
1. Барабой В.А. Чернобыль: через 10 лет. Медицинские
последствия радиационных катастроф / Под ред. Гродзинского Д.М., - К.:
Чорнобильінтернінформ, 1996. - 187 с.
2. Эйдус Л.Х. О механизмах инициации эффекта малых доз
// Радиац.
биол. Радиоэкол. - 1994. - Т. 34, вып. 6. - С. 748-758.
3. Zakrzhevskaya D. The effect of proteinase inhibitors on the level of degradation and unseheduled DNA synthesis in LIV- and g-irradiated cell
// Stud. Biophys. – 1990. – №1. – P. 31-40.
4. Кузин А.М. Стимулирующее действие ионизирующего
излучения на биологические процессы. М.: Атомиздат, 1977. - 136 с.
5. Volpe P., Parasassi
T., Sapora O., Ravagnan G., Eremenko T. Influence of doses of radiation on the
DNA double helix, gene expression and membranal state // International J.
Radiat. Medicine. – 1999. - 1(1). - P. 78-89.
6. Гераськин С.А. Концепция биологического действия малых
доз ионизирующей радиации на клетки // Радиац.
биол. Радиоэкол. – 1995. – Т. 35, №5. – С. 571-580.
7. Кристич Р.В. Гистологическая энциклопедия:
Иллюстрированная энциклопедия гистологии человека. – Берлин-Гейдельберг-Токио:
Из-во Шпрингера, 1984. – 341 с.