Биогеография,
биоресурсоведение, биоразнообразие
К.б.н.
Реуцкая В.В.
Российский
государственный социальный университет, Россия
Проблемы
восстановления генетического разнообразия в лесных экосистемах Среднерусской
Генетическое
разнообразие по существу характеризует
определённый уровень генотипических гетерозиготности, полиморфизм
популяций. Среди составных блоков биоразнообразия лесных экосистем генетическое разнообразие наименее изучено. В то
же время во многих случаях генетическая структура основных лесообразующих пород
может иметь определяющее значение для приспособления древесных пород к
разнообразным условиям произрастания. Гнетическое разнообразие является основой
коадаптации хозяинных и паразитических популяций, формирования устойчивых
биосоциальных отношений, основой естественного отбора, микроэволюционных
процессов, эволюции.
Исследования
проводились в крупнейших лесных массивах Среднерусской лесостепи – Шиповом лесу
и Усманском бору. Шипов лес является наиболее типичной дубравой подзоны
южной лесостепи. Высокими наследственными качествами отличаются и жёлуди
шиповского дуба.
По ландшафтному
районированию Шипов лес находиться на юго-восточной окраине лесостепной
провинции Среднерусской возвышенности, в подзоне южной лесостепи. Наиболее
распространёнными типами леса являются снытьевая дубрава (31,8 %) и
снытьево-осоковая дубрава (41,2
%). Снытьевая дубрава (Д2)
произрастает на высокоплодородных почвах плато или очень пологих склонах всех
экспозиций. Преобладают мощные чероноземы, темно-серыми и темно-коричневыми
суглинками на глубоких (до15-20 м) лессовидных отложениях. Бонитет Ia. В первом ярусе
преобладает дуб черешчатый с примесью ясеня и клена, липы и осины. Второй ярус
состоит из клена полевого, липы, ильма и яблони. Подлесок густой состоит из
лещины, клена полевого, реже бересклета европейского и бородавчатого. Травяной
покров хорошо развит. Преобладает сныть. Помимо, встречается копытень,
ясменник, звездчатка, реже сочевник, медуница, купена, осока волосистая.
Насаждения отличаются высоким уровнем композиционного разнообразия.
Снытьево-осоковая дубрава ( Д2-1) занимает
среднеплодородные почвы на пологих склонах разных экспозиций. Преобладают
среднемощные черноноземы, серые и коричневые суглинистые и глинистые на
глубоких лессовидных отложениях. Первый ярус представлен дубом (II) с примесью ясеня и клена.
Второй ярус – клен полевой, остролистный, ильм, липа, яблоня и груша. Подлесок
средней густоты, состоит из лещины, клена полевого, татарского, бересклета
европейского и бородавчатого. Травяной покров относительно хорошо развит, с
преобладанием сныти, осоки и звездчатки. Реже встречается ясменник, гравилат,
единично фиалка, будра, копытень.
Усманский бор является крупным
лесным массивом Среднерусской лесостепи, расположен между по левому берегу реки
Воронеж и ее притокам – Усмани, Ивнице. По расположению он входит в Левобережный
придолинно-террасный район подзоны типичной лесостепи лесостепной провинции
Окско-Донской низменности.
Чаще
других здесь встречаются дубняки осоковые, осоково-снытевые и
черемухово-снытевые. Первый ярус в них образован старовозрастным (до 150 лет)
дубом с небольшой примесью ясеня; во втором ярусе — клен остролистный, липа; в
третьем — обычен клен полевой. В подросте встречаются клен остролистный, дуб,
ясень, вяз, липа и другие породы, в густом подлеске — лещина, бересклет,
черемуха. Почва густо поросла снытью, осокой волосистой, звездчаткой, копытнем,
медуницей и другими характерными для дубрав травами.
История
освоения дубовых лесов Усманского бора сходна с историей освоения Шиповской
дубравы. И здесь в основном преобладают порослеввые насаждения с некоторым
включением семенных древостоев. Значительную часть насаждений составляют лесные
культуры.
Таким
образом, оба объекта – дубравы Шипова леса и дубравы Усманского бора имеют
определённую общность. Основное различие – географическое расположение (Шипов
лес является самым южным лесным массивом в Среднерусской лесостепи, у
Усманского бора – более северное положение) и тот факт, что Усманский бор
подвержен более сильной рекреационной нагрузке; санитарно-патологическое
состояние древостоев дуба в Усманском бору ниже, чем в Шиповом лесу.
Исследования проводились в осоко-снытьевых насаждениях обоих лесных массивов.
В таблице
1 представлены результаты сравнительной оценки аллельного и генного
разнообразия насаждений дуба черешчатого (Quercus robur) в Шиповом лесу и
Усманском бору.
Таблица 1 – Аллельное и генное разнообразие
насаждений Quercus robur в Шиповом лесу и Усманском бору
|
Исследуемый
локус |
Число
аллелей |
|
|
Шипов
лес |
Усманский
бор |
|
|
Phosphoglucose-Isomerase-B |
4 |
2 |
|
Isocitrat-Dehydrogenase-B |
3 |
3 |
|
Aminopeptidase-B |
7 |
3 |
|
Glutamai-Dehydrogenase |
3 |
3 |
|
6-Phosphogluconat-Dehydrogenase-B |
2 |
2 |
|
Saure
Phosphatase-A |
2 |
2 |
|
Saure
Phosphatase-C |
3 |
3 |
|
NADH-
Dehydrogenase-A |
3 |
3 |
|
NADH-
Dehydrogenase-B |
4 |
4 |
|
Menation-Reductase |
4 |
2 |
|
Phosphatglucomutase-A |
4 |
2 |
|
Aspartataminotransferase-B |
3 |
3 |
|
Генное
разнообразие ( ∑ ) |
42 |
32 |
|
Аллель/локус
(в среднем) |
3,5 |
2,7 |
Лабораторный
анализ проведен в отделе защиты леса Высшей лесной школы Эберсвальде (Eberswalde , FRG). Оценивались 10
энзимных систем 100 деревьев от каждого исследуемого насаждения.
Как
следует из таблицы 1, насаждения дуба черешчатого в Шиповом лесу отличаются
более высоким уровнем генетического разнообразия по сравнению с насаждениями
дуба черешчатого в Усманском бору (генное разнообразие 42, среднее число
аллелей 3,5 против генного разнообразия 32, среднего числа аллелей 2,7).
Интерпретация
результатов сравнительной оценки аллельное и генное разнообразия
насаждений дуба черешчатого в Шиповом
лесу и Усманском бору заключается в следующем: снижение генетического
разнообразия главной лесообразующей породы (дуба черешчатого) коррелирует cо
снижением уровня биорезистентности и общей стабильности насаждений дуба
черешчатого. Этот вывод соответствует литературным данным: возможно большее
генетическое разнообразие популяций лесообразующих пород является
предпосылкой экологической стабильности настоящих лесов и их будущих поколений (Stöcker, Rommerkirchen,
2002; Zaspel, 2000) [1], [2].
Сохранение генетических ресурсов имеет решающее значение для развития
биологического разнообразия. Эволюция биосферы, постоянно происходящая на
всех уровнях жизни, базируется прежде всего на генетических процессах, на
мутациях и рекомбинациях. В этом основа для приспособления, естественного
отбора в многообразных условиях жизни.
Снижение
генетического разнообразия главных лесообразующих пород неизбежно ведёт к
потере биорезистентности, общей стабильности устойчивого развития насаждений, к
снижению эффективности их защитных, экологических, экономических и социальных
функций.
Генетическая
инвентаризация, основанная на исследовании сравнительного состава изоэнзимов
генных локусов является наиболее эффективным современным методом получения
информации о генетической внутрипопуляционной изменчивости взрослых насаждений.
О
внутрипопуляционном разнообразии лесных древесных растений можно судить на
основе оценки коэффициента наследуемости в узком смысле. Определялась
жизнеспособность самосева сосны в очагах корневой губки (Heterobasidion annosum)
и материнских деревьев в прилегающей периферийной части очагов патогенна в
Усманском бору. Расчёт h2 проводился по
модифицированной нами формуле Шварца-Уирдена:
h2 = Vo / Vp ,
(1)
где h2 - коэффициент
наследуемости в узком смысле фактора жизнеспособности, Vo - средняя
жизнеспособность потомств (offspring),
Vp - средняя
жизнеспособность родительских деревьев (parents).
Результаты
оценки h2 устойчивости сосны обыкновенной к корневой
губке в Усманском бору представлены в таблице 2.
Как
следует из таблицы, уровень наследуемости жизнеспособности сосны в очагах
корневой губки в ювенильной стадии достаточно высок (h2 = 0,74). Этот уровень наследуемости позволяет прогнозироать эффективность
селекции сосны на устойчивость к корневой губке в насаждениях Усманского бора.
Таблица 2 – Наследуемость в узком смысле
жизнеспособности сосны (h2) в очагах корневой
губки
|
№№ проб |
n |
Vo |
n |
Vp |
|
1 |
15 |
2,1 |
35 |
2,1 |
|
1 |
18 |
2,0 |
28 |
3,0 |
|
2 |
24 |
2,2 |
34 |
2,2 |
|
3 |
16 |
1,8 |
36 |
2,8 |
|
4 |
14 |
1,6 |
24 |
3,6 |
|
5 |
12 |
2,4 |
32 |
2,4 |
|
6 |
21 |
2,3 |
31 |
2,3 |
|
7 |
23 |
1,7 |
33 |
2,7 |
|
8 |
16 |
1,6 |
36 |
3,6 |
|
9 |
15 |
1,9 |
15 |
2,9 |
|
10 |
11 |
2,7 |
23 |
3,7 |
|
11 |
10 |
2,8 |
18 |
2,8 |
|
12 |
18 |
2,0 |
18 |
3,0 |
|
13 |
14 |
2,1 |
24 |
2,1 |
|
14 |
16 |
2,3 |
26 |
3,3 |
|
15 |
8 |
3,1 |
18 |
3,1 |
|
16 |
12 |
2,8 |
16 |
3,8 |
|
|
∑
= 263 |
Ср.= 2,3 |
∑
= 447 |
Ср.= 3,1 |
|
h2 = 2,3 : 3,8 = 0,74 |
||||
|
Примечание.
Определеник Vo и Vp на уровне значимости α = 0,05 |
||||
С позиций генетического
разнообразия представляет интерес фенотипический признак «цвет семян» древесных
пород. Нами изучалась изменчивость по признаку "цвет семян" в
сосновых насаждениях Усманского бора в сравнении с Хреновским бором.
Были выделены две расы
сосны по цвету семян: чёрно-семенная и светлосеменная. Сбор семян - популяционный.
Результаты исследований представлены в таблице 3.
Таблица 3 - Сравнение
дисперсий S2 по цветосеменным расам
(чёрно-семенной и светлосеменной) сосны обыкновенной в монокультурах и
естественных насаждениях Хреновского и Усманского бора (a = 0,05; n1 = n2 = 500; возраст 80-90
лет)
|
Лесной
массив |
Характер
насаждений |
Критерий
Фишера |
||
|
Монокультуры |
Смешанный
лес |
Ffact |
Ftab |
|
|
Хреновской
бор |
16,40 |
32,62 |
1,99 |
1,56 |
|
Усманский
бор |
18,20 |
34,10 |
1,87 |
|
|
Критерий
Фишера |
1,11 |
1,05 |
|
|
Как следует из таблицы 3, дисперсия признака
"цветосеменные расы сосны обыкновенной" в природном лесу значительно
выше, чем в монокультурах, как в Хреновском бору, так и в Усманском. В то же
время различие дисперсий отдельно в пределах монокультур и отдельно в пределах
естественных насаждений между Хреновским бором и Усманском статистически
недостоверна.
Биологический смысл
результатов приведенной таблицы в том, что в искусственно созданных насаждениях
снижается феннотипическая изменчивость, определённую часть которой составляет
генотипическая изменчивость. Этот факт имеет негативное значение не только для
настоящего насаждения, но и особенно для будущих поколений лесообразующих
пород. Практический вывод заключается в том, что при создании насаждений следует
проектировать повышение генотипического, как составную часть фенотипического
разнообразия. При этом существенно учитывать хозяйственно ценные признаки
вводимых в насаждение биотипов древесных пород. Применительно к цветосеменным
расам сосны обыкновенной надо иметь в виду, что светлые семена обладают более
высокими абсолютными значениями хозяйственно ценных признаков.
В
заключение следует отметить, что генетическая составляющая общего
биологического разнообразия исключительно важна для устойчивого развития
насаждений, особенно с позиций паразито-хозяинных отношений. Флор (Flor, 1959, 1971) [3] первым
задался целью изучить генетику обоих компонентов системы
"хозяин-патоген", конкретно –
"Linum usitatissimum - Melampsora lini". Он пришёл к выводу о том, что
каждому гену, обусловливающему резистентность хозяинного растения,
соответствует ген патогена, обусловливающего вирулентность патогена. Другими
словами, существуют косвенное генетическое взаимодействие между хозяином и патогеном.
Пирсон (Person, 1959, 1962) [4] показал, что отношения между хозяином и
паразитом индуцируют генетический полиморфизм в популяциях обоих партнёров:
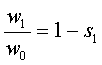
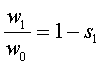
, (17)
где w1 – приспособленность хозяинного растения, w0 – приспособленность паразита, s1 - cелекционный коэффициент
хозяинного растения.
Оценка
этого соотношения на фенотипическом уровне при сравнения поражаемости дуба черешчатого (Quercus robur)
и распространением базидиокарпов его патогена – ложного дубового трутовика (Phellinus robustus)
представлено в таблице 4.
Таблица 4 –
Корреляционное отношение ( η ) между уровнями фенотипического
разнообразия Quercus robur ( DA
) и Phellinus robustus
(DPh)
|
Исследуемые параметры |
n число наблюдений |
η корреляционное
отношение |
m ошибка
корреляционного отношения |
α уровень
значимости, % |
|
DA ÷ DPh |
12 |
0,82 |
±0,06 |
5 |
Как следует из таблицы
4, корреляционное отношение (0,82) существенно при 5 % уровне значимости. Биологический смысл данного феномена во
взаимосвязанности биоразнообразия компонентов паразито-хозяинной системы "древесное
растение – паразит».
Таким образом, уровень генетического разнообразия дубовых
насаждений коррелирует с уровнем их биорезистентности и общей стабильности;
возможно большее генетическое разнообразие популяций лесообразующих пород
является предпосылкой экологической стабильности настоящих лесов и их будущих поколений; сохранение
генетических ресурсов имеет решающее значение для развития биологического разнообразия. Уровень наследуемости жизнеспособности сосны в
очагах корневой губки в ювенильной стадии достаточно высок (h2
=
0,74) позволяет прогнозироать эффективность селекции сосны на устойчивость к
корневой губке в сосновых насаждениях Усманского бора. Корреляционное отношение
между уровнями фенотипического разнообразия
Quercus robur и его патогенном Phellinus robustus проявляется на уровне 0,82.
Литература:
1. Stöcker,
G. Beiträge zur Strukturanalyse natürlicher und forstlich bestimmter
Fichten-Ökosysteme im Nationalpark Hochharz / G. Stöcker, A.
Rommerskirchen //
Beiträge für Forstwirtschaft und Landschaftsökologie. 1/2002. Band 36. 2002. – S. 6 – 13.
2. Zaspel I. Waldschäden und genetische Strukturen in
Beständen einheimischer Eichenarten / I. Zaspel, H. Hertel, T. Stauber //
Beiträge für Forstwirtschaft und Landschaftsökologie, 2002, N.
3. – S. 111 – 115.
3. Flor H. H. Inheritance of
pathogenicity in Melampsora lini / H. H. Flor //
Phytopathology, 32, 1942. – P. 653-669.
4.Person C. The gene-for-gene concept
/ C. Person, D. J. Samborski and R.
Rohringer // Nature 194, 1962. – P. 561
– 562.