И.И. Дмитриев, г. Горно-Алтайск, E-mail: dmitrievilya86@gmail.com;
А.В. Бондаренко, доктор
биол. наук, доц. ГАГУ, г. Горно-Алтайск, E-mail: gf@gasu.ru
А.Г. Манеев, канд. Биол.
Наук, доц. ГАГУ, г. Горно-Алтайск, E-mail: gf@gasu.ru
Население дневных
бабочек (LEPIDOPTERA, DIURNA)
Юго-Западной Тувы и
сопредельных территорий
Оценка биологического разнообразия и выявление структурных особенностей сообществ
– одно из фундаментальных направлений современных биогеографических
исследований. Трудоемкость таких работ общеизвестна, поэтому, по крайней мере,
на первых этапах, целесообразно проводить исследования отдельных
таксономических групп организмов [1]. В этом отношении дневные чешуекрылые, как
широко распространенные, способные к перелетам и очень чувствительные к
изменениям среды насекомые, весьма перспективны.
Материалы и методы исследования. В период с 2006 по 2009 гг. на
территории Юго-Западной Тувы, Юго-Восточного Алтая и Северо-Западной Монголии авторамиом
пройдено 430 км и обследовано 73 местообитания, что составило172 часа маршрутных
учетов. Кроме того, для выявления пространственной
структуры и организации населения дневных бабочек Юго-Западной Тувы и
сопредельных территорий были использованы материалы А.В. Бондаренко по
Юго-Восточному Алтаю [2] - 63 местообитания, Центральной Туве - 10 и
Северо-Западной Монголии [3] – 37 местообитаний. В результате полученные данные
в общем объеме 183 местообитания,
позволили выявить структуру населения дневных бабочек исследуемой территории.
Учеты и отлов дневных
бабочек проводились автором по методике А.П. Кузякина [4]. Для оценки плотности
использовалась методика учета на трансектах [5; ,6],
уточненная Ю.П. Малковым [7]. Данные по одинаковым местообитаниям в пределах
одних и тех же поясов усреднены. Для выявления особенностей пространственной
организации использованы оценки сходства населения по коэффициентам Жаккара [8]
в модификации Р.Л. Наумова [9]. На их основе
проведен кластерный анализ методом факторной классификации с расчетом внутри- и
межклассового сходства населения и доразбивкой крупных классов [10]. Материалы
обработаны с помощью программного обеспечения банка данных лаборатории
зоологического мониторинга ИСиЭЖ СО РАН (г. Новосибирск).
Классификация
населения. В результате кластерного
анализа выделено 3 типа населения, каждый из которых
разделяется на подтипы (8 подтипов). В большинстве подтипов были выделены
классы, что связано с разнообразием вошедших в них выделов. Деление на типы
совпадает с различием в региональной принадлежности, а на подтипы с характером
растительности (степенью облесенности) и увлажнением.
Пространственно-типологическая
структура населения дневных бабочек Юго-Западной Тувы и сопредельных
территорий. Под
пространственно-типологической структурой животного населения понимается общий
характер его территориальных изменений, выявленных по морфологическому сходству
сообществ (их облику), без учета их сопряженности на местности [11]. Она
отражается структурным графом. Структурный
граф (рис. 1) построен на уровне классов населения при пороге значимости выше 9
единиц. На схеме вертикальное расположение иллюстрирует изменение облика
населения дневных бабочек по мере смещения с севера на юг, а горизонтальное - с
запада на восток. В целом, на графе отчетливо прослеживается три сгущения
связанные с региональностью. В первое из них входят сообщества, преимущественно,
Юго-Восточного Алтая и отчасти Юго-Западной Тувы, через которое осуществляется
связь со второй группой сообществ. Второе сгущение образовано классами
населения дневных бабочек Тувы, третье – Северо-Западной Монголии. Расположение
групп соответствует географическому расположению изученных территорий. Внутри
каждой из провинций прослеживаются изменения распределения дневных бабочек в
зависимости от облесенности местности. Так,
облесенность усиливается с востока на запад отдельно внутри каждой из
провинций.
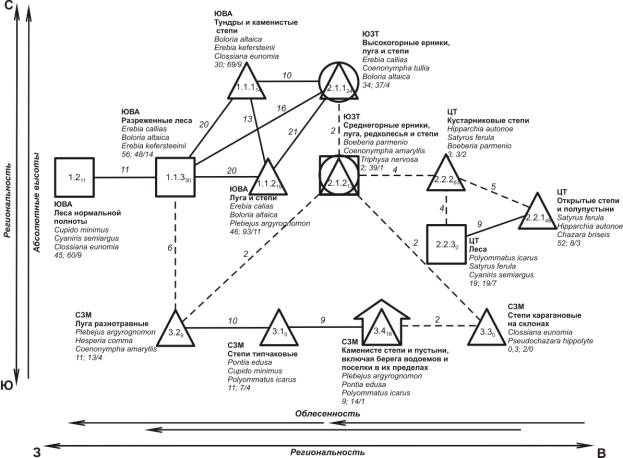
Рис. 1. Пространственно-типологическая структура населения дневных
бабочек Юго-Западной Тувы и сопредельных территорий:
Условные
обозначения: треугольником обозначается фауна открытых обедненных
по продуктивности местообитаний, кругом – мозаичных, квадратом – облесенных,
значком дома – местообитания под воздействием антропогенного фактора. Внутри значков представлены номера таксонов
соответствующей классификации, индексы характеризуют внутригрупповое сходство,
непрерывные линии между значками означают сверхпороговое сходство, прерывистые
– максимальные значения меньше порога. Около линий связи приведены значения
межгруппового сходства, около значков – плотность населения особей/га; общее количество
встреченных видов в пересчете на 1 вариант населения/число фоновых видов.
Стрелки около перечня основных структурообразующих факторов среды указывают
направление увеличения их влияния и тренды в населении.
Абсолютные высоты местности также играют
важную роль в распространении и неоднородности населения бабочек, это отражено
в расположении таксонов по вертикали. Население бабочек Северо-Западной
Монголии образует горизонтальный ряд, так как все местообитанияместообитания,выделы,
вошедшие в таксоны 3.1 – 3.4, находятся на,
примерно, одинаковой высоте над уровнем моря.
Плотность населения максимальна в
Юго-Восточном Алтае (классы 1.1.1 – 1.1.3, 1.2). Здесь значения колеблются от 30 до 56 особей/га. Столь же
высокие показатели свойственны высокогорным ерникам, лугам и степям
Юго-Западной Тува (класс 2.1.1) и открытым степям и полупустыням Центральной её
части (класс 2.2.1). В остальных случаях плотность населения значительно ниже и
чаще всего не превышает двадцати особей/га. Следует отметить, что в карагановых
степях на склонах гор в Северо-Западной Монголии (класс 3.3) суммарное обилие
составляет 0,3 ос./га. Это очень низкий показатель даже
для суровых условий Северо-Западной Монголии. Объясняется это тем, что эти
кустарники произрастают преимущественно на теневых склонах отличающихся
недостаточной теплообеспеченностью и сниженной продуктивностью биоценозов.
Максимальные показатели видового
разнообразия свойственны лугам и степям Юго-Восточного Алтая (класс 1.1.2). В
лесах, тундрах и каменистых степях Юго-Восточного Алтая, отмечено 60 и 69 видов,
соответственно. В разреженных лесах Юго-Восточного Алтая (класс
1.1.3) зарегистрировано 48 видов дневных бабочек. Столь же высокие показатели
свойственны открытым степям и полупустыням Центральной Тувы (класс 2.2.1) и в
Юго-Западной Туве (2.1.1 и 2.1.2). В остальных случаях количество видов не
превышает и двадцати видов. Таким образомобразом,
в Юго-Восточном Алтае отмечено от 48 до 93 видов. В населении Юго-Восточного Алтая также больше и фоновых видов (от
9 до 14), в других случаях их значительно меньше.
Сверхпороговые связи между таксонами
отмечены лишь внутри провинций и ниже между ними. Информативность структурных
представлений на уровне классов составляет 34% учтенной дисперсии.
Итак, территориальная неоднородность
населения булавоусых чешуекрылых исследуемой территории на уровне классов в
наибольшей степени зависит от региональности, увлажнения и абсолютных высот
местности. В целом, плотность населения булавоусых
чешуекрылых в Юго-Восточном Алтае несколько выше, чем в Туве и особенно в
Северо-Западной Монголии. Это связано с низкой продуктивностью биоценозов Северо-Западной Монголии. То же соотношение
прослеживается и в показателях как видового богатства, так и количества фоновых
видов. В результате Северо-Западную Монголию можно считать территорией, условно
субоптимальной для дневных бабочек. Высокие показатели плотности населения,
видового разнообразия, количества фоновых видов в Юго-Восточном Алтае можно
объяснить достаточно высокой продуктивностью биоценозов, разнообразием
растительных сообществ, ии, следовательноследовательно,
и достаточным количеством экологических ниш.
Региональность в данном случае
определяется отличиями климата, растительности и характером хозяйственной деятельности.
В целом, у дневных бабочек пространственно-типологическая структура населения
проще, чем у птиц и млекопитающих, за счет миграционного выравнивания [12].
Структурообразующие
факторы. Проведенные расчеты с
помощью линейной качественной аппроксимации (качественного аналога
регрессионной модели [13]) показали, что наибольшее влияние на дифференциацию
населения булавоусых чешуекрылых оказывает региональность (27 % учтенной
дисперсии матрицы коэффициентов сходства). Значительно меньшее воздействие оказывает
увлажнение (10%) и абсолютные высоты местности (7,8%). Влияние облесенности на
организацию населения дневных бабочек оценивается лишь в 1,3% учтенной
дисперсии. По результатам расчетов наименьшее влияние оказывает характер
рельефа (в том числе крутизна склонов) – 0,1%.
В результате можно сделать вывод, что
наибольшее влияние на дифференциацию населения дневных бабочек Юго-Западной
Тувы и сопредельных территорий оказывает региональность, абсолютные высоты и
увлажнение. Набор выявленных факторов можно считать достаточно полным, так как
информативность представлений о структурообразующих факторах среды составляет
34% учтенной дисперсии.
Библиографический список
1. Равкин, Ю.С. Лукьянова, И.В. География
позвоночных южной тайги Западной Сибири. Новосибирск: Наука, СО, 1976.
2. Бондаренко, А.В.
Зоогеография булавоусых чешуекрылых Юго-Восточного Алтая. - Томск: Изд-во Томского университета, 2005
3. Бондаренко, А.В.
Сравнительная характеристика населения булавоусых чешуекрылых (Lepidoptera, Rhopalocera) Тывинской
и Убсу-Нурской котловин // Вестник Томского государственного университета.
Бюллетень оперативной научной информации «Оценка биоресурсов трансграничной
биосферной территории (ТБТ): Россия, Монголия, Казахстан, Китай». - 2006. №107.
- Часть 1.
4. Кузякин, А.П.
Зоогеография СССР // Уч. зап. Моск. пед. ин-та им. Н.К. Крупской. – М. - 1962. - Т. 109. - Вып. 1.
5. Yamomoto, M. Notes on the methods of belt transect census of
butterfies // J. Fac. Sci. Hokkaido Univ. - 1975. - Vol. 20.- № 1. - Ser. V1.
6. Pollard, E. A
method for assessing changes in the abundance of butterflies // Biol. Coserrv. - 1977. - Vol. 12.
- № 2.
7.
Малков, Ю.П. К методике учета дневных бабочек // Животный мир Алтае-Саянской
горной страны - Горно-Алтайск, 1994.
8. Jaccard, Р. Lois de distribution fiorale danse la zone alpine // Bull. Soc. Vaund. Sci.
Nat., 1902.
9. Наумов, Р.Л. Птицы природного очага
клещевого энцефалита Красноярского края: автореф. дис. … канд. биол. наук,
Москва, МГУ, 1964.
10. Трофимов, В.А. Модели и методы качественного
факторного анализа матрицы связи, Проблемы анализа дискретной информации,
Новосибирск: Наука СО, 1976.
11. Равкин, Ю.С.
Пространственная организация населения птиц лесной зоны (Западная и Средняя
Сибирь). - Новосибирск: Наука, 1984
12. Равкин, Ю.С. Особенности
биоразнообразия Российского Алтая на примере модельных групп животных / Ю.С.
Равкин, С.М. Цыбулин, С.Г. Ливанов [и др.]
// Успехи современной биологии. - 2003. - Том 123. - №4.
13. Равкин, Ю.С. Птицы
лесной зоны Приобья / Ю.С Равкин, В.А. Куперштох, В.А.Трофимов. - Новосибирск: Наука, 1978.