УДК 581.1
Динамика активности
сахарозорасщепляющих ферментов у растений карельской березы в течение
вегетационного периода.
Мощенская Ю.Л., Галибина Н.А.
Учреждение Российской академии наук Институт леса
Карельского научного центра РАН, Петрозаводск.
Известно, что в
растительном организме значительная часть углеводов, образующихся в процессе
фотосинтеза, расходуется при окислении в процессе дыхания (с высвобождением
энергии необходимой для роста), а также откладывается в виде запасных
метаболитов. Основная же масса метаболитов используется непосредственно в
акцепторных тканях, где они служат субстратом для синтеза полисахаридов и
других компонентов клеточных стенок (целлюлозы, гемицеллюлоз, пектина,
лигнина). [1]. Утилизация сахарозы в акцепторных тканях обеспечивает
поддержание относительно невысокого уровня сахарозы в ситовидных трубках,
необходимого для нормального флоэмного транспорта.
В
Институте леса КарНЦ РАН в исследованиях, проведенных на деревьях березы
повислой Betula pendula var. pendula с
нормальным строением тканей ствола и B. pendula var. carelica с
узорчатой древесиной, было показано, что образование узорчатой древесины
карельской березы индуцируется избыточным накоплением сахаров в проводящей
флоэме [2, 3].
Вовлечение сахарозы в метаболизм растения
осуществляется благодаря ее ферментативному расщеплению с участием инвертазы (ЕС 3.2.1.26) или сахарозосинтазы (ЕС 2.4.1.13). В зависимости от того,
активность какого из этих двух ферментов будет преобладать в акцепторной ткани,
будет меняться направленность метаболизма сахарозы в растении. Так, в
результате сахарозосинтазного расщепления сахарозы, образуется УДФ-глюкоза,
идущая на синтез структурных компонентов клеточной стенки [7], а также
на синтез крахмала [4, 5] и каллозы [8], в то время как расщепление сахарозы с
участием инвертазы приводит к образованию гексоз, индуцирующих образование
запасных метаболитов, а также обеспечивающих поддержание тургора и рост клеток
растяжением [6]. При этом
наиболее значимой является инвертаза клеточной стенки, поскольку включение
инвертазы в процесс транспорта сахарозы происходит в том случае, если
плазмодесменные связи нарушаются, и перенос сахарозы происходит апопластически
[6]. Поэтому целью нашего исследования
было определение активности сахарозосинтазы и апопластной инвертазы, а также
изучение соотношения инвертазного и сахарозосинтазного пути включения сахарозы
в метаболизм у растений карельской березы с разной структурой древесины.
Исследования
проводились на 40-летних растениях карельской березы с проявившимися и
непроявившимися признаками узорчатости ствола. Отбор образцов производили в
течение вегетационного периода (май, июнь, июль, октябрь).
У
безузорчатых растений в мае, когда основным акцептором являются листья,
во флоэме наблюдается высокая активность апопластной инвертазы. В ксилеме очень
низкую активность проявляет инвертаза клеточной стенки (ApInv), сахарозосинтаза (SS)
в этот период активность не проявляет (рис.1). В июне, в период интенсивных
ростовых процессов во флоэме происходит снижение активности апопластной
инвертазы. Активность сахарозосинтазы во флоэме остается низкой в течение всего
вегетационного периода. В ксилеме к июню происходит резкое увеличение
активности сахарозосинтазы (рис.1.), что связано с ее участием в процессах
синтеза компонентов клеточной стенки в период формирования тканей ксилемы.
Июльская точка отбора проб характеризуется отсутствием дождей и высокими
температурами, поэтому, в нашем исследовании июль - период преждевременного
торможения камбиальной деятельности. В этот период наблюдается резкое снижение
активности сахарозосинтазы и отсутствие активности апопластной инвертазы в
ксилеме. Во флоэме в июле возрастает активность сахарозосинтазы, что может быть
вызвано синтезом запасных нерастворимых углеводов. Так в этот 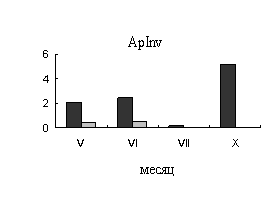 период, во
флоэме происходит накопление крахмала (рис.2.).
период, во
флоэме происходит накопление крахмала (рис.2.).
Б
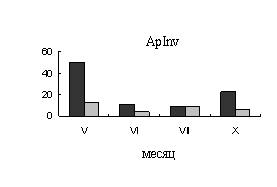
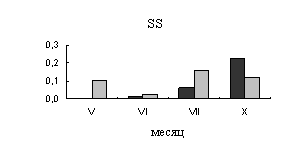
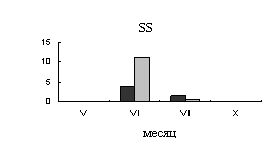
Рис.1. Активность
ферментов метаболизации сахарозы во флоэме (А) и ксилеме (Б) 40-летних деревьев
карельской березы (темные столбики – с проявившейся узорчатостью, светлые
столбики – безузорчатые растения). По оси ординат – активность ферментов (ApInv-мкмоль распавшейся сахарозы/г сырой
ткани, SS – мкмоль распавшейся сахарозы/г белка).
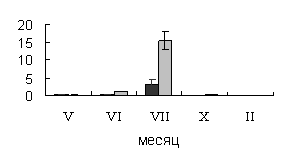
Рис.2.
Динамика содержания крахмала в тканях флоэмы 40-летних деревьев
карельской березы (темные столбики – с проявившейся узорчатостью, светлые
столбики – безузорчатые растения). По оси ординат – содержания крахмала (мг/г
сырой ткани).
А
В октябре в
ксилеме происходит снижение активности ферментов и их активность близка к нулю.
У
узорчатых растений карельской березы также наибольшую активность
проявляет апопластная инвертаза. В тканях флоэмы ее значения значительно
превышают таковые у безузорчатых деревьев. В ксилеме узорчатых растений
активность сахарозосинтазы значительно ниже, чем у безузорчатых. Как и у
безузорчатых растений максимальная активность сахарозосинтазы узорчатых
растений наблюдается в июне, а к июлю снижается, хотя и не столь резко как у
безузорчатых. Следует отметить, что у растений с узорчатой структурой древесины
апопластная инвертаза проявляет высокую активность и в тканях ксилемы и
достигает наибольшего значения в октябре. Возможно, это связано с тем, что у
деревьев карельской березы в ксилеме присутствуют аномальные прослойки
паренхимы, которые дают возможность дополнительного запасания метаболитов в
период подготовки растения к покою.
Изучение
активности ферментов метаболизации сахарозы в тканях ствола карельской березы с
разной степенью узорчатости ствола показало, что ткани карельской березы
отличаются высокими значениями апопластной инвертазы, и только в июне в период
активного роста ксилемы, в этой ткани преобладает сахарозосинтазная активность.
Активность апопластной инвертазы узорчатых растений была выше в течение всего
вегетационного периода. Так, образование гексоз в результате инвертазного
расщепления сахарозы вызывает активизацию синтеза запасных метаболитов и превращение камбиальных производных в клетки
запасающей паренхимы, которые являются структурной основой узорчатой ткани
карельской березы.
Список
литературы:
1.
Веретенников
А.В. Физиология растений. Москва, Академический проект,
2006. 479 с.
2. Галибина Н.А., Новицкая Л.Л., Софронова И.Н. Динамика сахаров в тканях ствола Betula pendula (Betulaceae) при выходе из зимнего покоя. // Растительные ресурсы. 2012. Т. 48. №4. С. 554 – 564.
3.
Новицкая Л.Л. Карельская береза: механизмы роста и
развития структурных аномалий. Петрозаводск: Verso, 2008. 144 p.
4. Asano T., Kunieda N., Omura Yu., Ibe H., Kawasaki T.,
Takano M., Sato M., Furuhashi H., Mujin T., Takaiwa F., Wu Ch., Tada Yu.,
Satozawa T., Sakamoto M., Shimada H. Rice SPK, a calmodulin-like domain protein
kinase, is required for storage product accumulation during seed development:
Phosphorylation of sucrose synthase is a possible factor. Plant Cell. 2002. V. 14(3). P. 619-628.
5. Kladnik A., Chamusco K., Chourey P. S., Dermastia M. In situ detection of programmed cell death in the
maize caryopsis // Periodicum
Biologorum. 2005.
V. 107. P. 11–16.
6. Koch K.E., Zeng Y. Molecular approaches to altered C partitioning: genes for sucrose metabolism//
J. Amer. Soc. Hort. Sci. 2002. V. 127. P. 474-483.
7.
Ruan, Y.L., Llewellyn, D.J., Furbank, R.T. Suppression of sucrose synthase gene
expression represses cotton fiber cell initiation, elongation, and seed
development // Plant Cell Online. 2003. V. 15. P. 952–964.
8. Salnikov VV,
Grimson MJ, Seagull RW, Haigler CH Localization of sucrose synthase and callose
in freeze-substituted secondary-wall-stage cotton fibers // Protoplasma 2003. V.
221. P. 175–184.