Биологическикие науки/ 2. Структурная ботаника и
биохимия растений
К.б.н.
Кирильчук Е.С.
Сумский национальный аграрный
университет, Украина
Онтогенетическая
структура популяций Trifolium repens на
пойменных лугах лесостепной зоны (Украина) в условиях
хозяйственного
пользования
Особи одной популяции различаются
по многим признакам: по календарному или биологическому возрасту (возрастные или
онтогенетические состояния), виталитету, размеру, соотношению особей двух полов
(в случае двудомных растений), наличию или отсутствию генетического
полиморфизма и другим особенностям. Анализируя их, можно судить о популяции в
данный момент времени, охарактеризовать ее прошлые и будущие состояния.
Одним из основных методов изучения особенностей
функционирования популяций в тех или иных условиях обитания является исследование
их онтогенетической структуры. Теоретические основы ее изучения были заложены
российской школой геоботаники: подразделение онтогенеза у растений на этапы
было предложено Т.А.Работновым [4] и позже дополнено А.А.Урановым [5]. Так,
онтогенез травянистых многолетних растений подразделяют на четыре периода,
каждый из которых включает те или иные возрастные состояния: латентный
(семена), предгенеративный (проросток, ювенильное, имматурное и виргинильное), генеративный
(молодое генеративное, среднегенеративное и старое генеративное) и
постгенеративный периоды (субсенильное, сенильное и отмершие растения). Определенный
этап онтогенеза характеризуется набором качественных признаков, критерии
оценивания которых, с одной стороны, сходны для всех видов растений (поскольку представляют
одинаковую схему, отражающую стадии развития), а с другой, – индивидуальны для каждого
вида в частности.
Особой индивидуальностью отличаются подходы к
определению качественных признаков для тех растений, у которых счетной единицей
является рамет.
Бобовый компонент в составе луговых травостоев
является основным источником протеина в кормовом сене. Одним из неотъемлемых видов
растений луговых фитоценозов является T. repens,
который, несмотря на низкорослость и малую доступность для выпасаемых животных,
составляет основу кормового потенциала лугов, используемых в качестве пастбищ. T. repens – луговое
вегетативно-подвижное растение, формирующее клоны. Счетной единицей при анализе
онтогенетической структуры его популяций является рамет – относительно
независимый элемент клона, являющийся результатом вегетативного размножения
генета. М.М.Паленова [3] предлагала оценку возрастного состояния
раметов (v1, v2, v3, g1, g2,
g3) проводить, опираясь на длину междоузлий и высоту листа. Л.Н. Денисова [1] определяла рамет T. repens как верхушечную меристему
со сформировавшейся осью побега.
С целью исследования онтогенетической структуры
популяций Trifolium repens на
пойменных лугах Лесостепи Украины (река Псел) в условиях выпаса (градиент пастбищной
дигрессии включает 5 ступеней от контрольного участка (КУ) до участка с
интенсивным выпасом (ПД4), разработаны
критерии оценки возрастных состояний T. repens,
а также проведен учет возрастных состояний в пределах его популяций и
проанализированы онтогенетические спектры.
На основании анализа морфогенеза T. repens мы предлагаем оценивать онтогенетический состав популяций,
основываясь на структуре отдельного рамета, который является самостоятельной
биологической единицей. В ходе исследования учитывались все узлы ползучего
побега, которые имеют зачаток листка или соцветия, или сформировавшийся листок
и соцветие, или и листок, и соцветие одновременно. При оценке возрастного
состояния раметов отмечалось их положение на побеге с его апикального конца
(рис. 1). Поскольку ведущий способ размножения T. repens – вегетативный, то особей семенного размножения
(генетов), по сравнению с особями вегетативного размножения (раметов), намного меньше.
Поэтому учет состояния раметов в полной мере отражает онтогенетическую
дифференциацию в популяции этого вида.
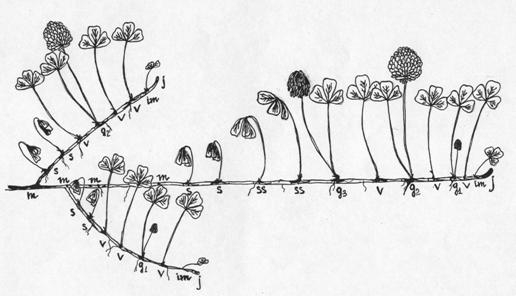
Рис. 1. Периодизация онтогенеза
T. repens
Предложенная периодизация онтогенеза T. repens приведена ниже:
Проростки (p) - всходы, семядоли 3-
Ювенильные растения (j) – появляется первый широко-округлый
листок, следующие - тройчастые. Появляются корни других порядков.
По отношению
к раметам используется следующая периодизация:
Ювенильные
растения (j) - начало появления листка из почки.
Імматурные (im) - листок будущего рамета полностью не
сформирован.
Виргинильные (v) – полностью сформированный рамет с одним
листком и придаточными корнями. Цветение отсутствует.
Молодые генеративные (g1) – начало появления у
рамета бутона.
Середние генеративные (g2) – цветущий рамет.
Рамет представлен одним хорошо развитым листком и соцветием.
Старые генеративные (g3) - листок подсыхает, соцветие
в фазе начала плодоношения.
Субсенильные (ss) - рамет засыхает, плодоношение завершается.
Сенильные (s) – полностью засохшие раметы.
Погибшие
раметы (m) – узлы на побеге, которые в прошлые вегетационные периоды характеризовались
наличием рамет.
Исследования показали, что на контрольных участках для T. repens в
пойме р. Псел характерны нормальные левосторонние возрастные спектры з преобладанием
догенеративных особей (виргинильных), спектры в отдельные
годы отличаются неполночленностью. Наиболее характерно выпадение возрастного
состояния g3. Индекс генеративности
[2] (отношение особей генеративного состояния к общему количеству особей в
популяции, выраженному в процентах) популяций T. repens на
контрольных участках невысокий - в среднем 8 %; индекс возобновляемости (отношение догенеративных особей к общему
количеству особей в популяции, выраженному в процентах) составлял 73 %; индекс старения (отношение особей
постгенеративного возрастного состояния к общему количеству особей в популяции,
выраженному в процентах) - 19 %. Такие типы спектра достаточно характерны
для T. repens, они и ранее регистрировались в условиях лугов лесной
зоны [3]. Отдельные авторы считают, что такой тип спектра - результат активного
вегетативного размножения [1]. Возрастная категория популяций, по Т.А.Работнову,
- инвазионная.
В
условиях нарастания пастбищных нагрузок для T. repens в пойме
р. Псел характерно сохранение характера онтогенетического спектра. Пик численности, как и на контрольных участках, на
пасквальном градиенте приходился на виргинильные особи. На выпасаемых участках
в 2-4 раза повышается доля субсенильных и сенильных раметов. Однако онтогенетический спектр приближается к
нормальному, он полночленный, хотя за счет ускорения темпов развития генеративных
раметов из популяций выпадают особи возрастного состояния g3. На
пасквальном градиенте у T. repens
выражена тенденция к снижению индекса возобновляемости (рис. 2).

Рис.
2. Общие тенденции изменения индексов возобновляемости (1) и старения (2) популяций
T. repens на пастбищном градиенте.
Выявленные
изменения популяционного статуса T. repens свидетельствуют о том, что интенсивный выпас оказывает отрицательное
влияние, усложняя семенное размножение и разрушая ниши возобновления его
популяций. В результате на пойменных лугах реки Псел под действием выпаса
происходит возрастание индексов старения популяций T. repens, а это,
безусловно, снижает их устойчивость на лугах при использовании их в качестве
пастбищ.
Литература:
1.
Денисова Л.Н. Пространственная и возрастная
структура популяций Trifolium repens (Fabaceae) в различных местообитаниях //
Ботан. журн., 1995. – Т. 80, № 5. – С. 18 – 25.
2.
Коваленко І.М. Структура популяцій основних
домінантів трав’яно-чагарничкового яруса в лісових масивах
Деснянсько-Старогутського національного парку - Автореф. дис... канд. біол. н.
- Київ, 2003. - 20 с.
3.
Паленова М.М. Отногенез и
структура ценопопуляционных локусов клевера ползучего // Сб. «Популяц. экология
растений». - М.: Наука, 1987. - С. 92-95.
4. Работнов
Т.А. Вопросы
изучения состава популяций для целей фитоценологии. // Проблемы ботаники -
М.-Л.: Изд-во АН СССР, 1950 - С. 465-483.
5. Уранов А.А. Возрастной спектр
фитоценопопуляций как функция времени и энергетических волновых процессов //
Биол. науки, 1975. - № 2. – С. 7 – 33.