Кривая амплитуды сокращений
лимфатических узлов у крыс
|
|
|
Подчелюстные Подмышечные Сердечные в Кишечные " Брыжеечные Почечные Печеночные Паховые |
Кривая частоты сокращений лимфатических узлов у крыс
Мырзаханова М.Н.
Карагандинский государственный
университет имени Е.А.Букетова
Становление сократительной
деятельности лимфатических узлов в
постнатальном онтогенезе.
Согласно исследованиям последних лет
(Ю.И.Бородин,2006) лимфатическая система
млекопитающих представлена прелимфатическими и лимфатическими
капиллярами, внутриорганными лимфатическими сосудами, отводящими афферентными
лимфатическими сосудами, лимфатическими узлами, отводящими эфферентными лимфатическими сосудами, грудным и
правым лимфатическими протоками.
К
настоящему
времени, хорошо изучены тонкая структурно-топографическое
строение и иннервация аппарата лимфообращения, установлены основные функций
(дренажная, иммунологическая, белоктранспортная и.т.п.) лимфатической системы, которые обеспечивают сохранение эндоэкологического гомеостаза внутренних сред организма.
Известно, что от каждого органа
или части тела отходят
несколько лимфатических сосудов, которые
на своём пути следования прерываются лимфатическими узлами. Количество лимфатических
узлов у представителей различных видов млекопитающих широко варьирует. Так, их
количество у свиньи составляет – 190, у крупного рогатого ската-300, у мелкого рогатого
скота-145 и у лошадей-8000. Средние размеры узлов колеблются от 0,9 до
10 см, реже до 20 см.
Впервые Мырзахановым
Н.М. (1987,2006,2007) установлено значение лимфатических узлов в транспорте
жидкостей, который в опытах in vitro зарегистрировал ритмическое
спонтанное сокращение лимфатических узлов тела (сомы) и внутренних органов, изучил иннервацию узла, влияние на них биологически активных веществ (норадреналин, ацетилхолин,
гепарин, гистамин, АТФ) у ряда
млекопитающих животных (крыса, овца, корова, свинья) и выдвинул концепцию о сосудисто-узловой системе
транспорта лимфы в животном организме (1995),которые нашли экспериментальное
подтверждение в опытах Р.С.Орлова (2001), Л.Э.Булебаевой (2001) и
М.Р.Хантурина (2002).
Разработка научных проблем о генезе
спонтанной ритмической сократительной функций лимфатических узлов, как и генез
самой лимфатической системы является мало изученным аспектом в учении о
лимфатической системе. Согласно одной теории лимфатические сосуды произрастают
из стенок вен, тогда как другая теория придерживается мнения о независимом от
вен, мезенхимальном происхождений элементов лимфатической системы, а что
касается вопроса о постнатальном становлений спонтанной ритмической
сократительной активности лимфатических узлов и сосудов вопрос остаётся
открытой. Известно что лимфатические узлы у плодов крупного рогатого скота и
овец закладываются уже к третьему месяцу утробной жизни (Иосифов,1942; Григорьев,1975). С возрастом узлы
уплотняются и утолщаются их соединительнотканные и гладкомышечные элементы. Внутренняя
структура лимфатических узлов зависит от их локализации: в области сомы, в
брюшной и грудной полостях (Славензон,1956; Кривский,1967). По данным
Н.Г.Игнатьева (1975) ягнята рождаются с функционально сформировавшейся
симпатической иннервацией лимфатических узлов, что чётко выявлялось при анализе
содержания катехоламинов в узлах в различные сроки постнатального периода.
Функциями лимфатических узлов
являются: лимфоцитопоэз, барьерно-фильтрационная, транспортная, иммунопоэз, обменная,
депонирование (в частности витаминов А и В), внутренней секреции (гепарин), разрушение
эритроцитов. Естественно предположит, что в выполнений выше перечисленных функций лимфатических
узлов немаловажное значение имеет изучение становления спонтанной ритмической
сократительной функций лимфатических узлов, в частности в постнатальном
онтогенезе.
Объектом исследования служили
изолированные, одиночные соматические (подчелюстной, подмышечный, паховый) и
висцеральные (сердечный, кишечный, брыжеечный, почечный, печеночный)
лимфатические узлы лабораторных крыс.
Изометрическое напряжение препаратов
регистрировали при помощи высоко – чувствительных механотронов типа 6 МХ-1Б
(Ерофеев и.др;1975) на чернильно пишущем самописце Н-327-5. Термостатирование
производили с помощью ультратермостата УТ-15
с точностью не ниже 0,2оС.
Спонтанные ритмические сокращения лимфатических узлов производили на 1, 5,
10, 15, 20 и 30 дни после рождения крысят. Анализ лимфонодулограммы производили
по общей картине кривой относительно изометрической линий, а также по частоте и
амплитуде сокращений узла, включающей 5
записей по одной минуте в течение 5 минут. Кроме того, с помощью курвиметра
определяли продолжительность кривой за 5
минут, которая служила характеристикой общего уровня биоэлектрической
активности узла (Собакин,1971).
Результаты исследований показали, что
характер спонтанной ритмической сократительной деятельности лимфатических узлов
имеют чёткие отличия в зависимости от сроков постнатального онтогенеза органа
(табл.1-2)
Таблица №1
Частота спонтанных сокращений лимфатических узлов у крысят в онтогенезе
|
Лимфатические
узлы |
период
наблюдений, дни |
|||||
|
1 |
5 |
10 |
15 |
20 |
30 |
|
|
Подчелюстные |
0,64±0,047 |
0,93±0,101 |
1,75±0,207 |
2,33±0,351 |
2,29±0,343 |
2,04±0,280 |
|
Подмышечные |
0,67±0,036 |
1,02±0,079 |
1,71±0,195 |
2,29±0,332 |
2,41±0,367 |
2,25±0,323 |
|
Сердечные |
0,23±0,080 |
0,27±0,006 |
0,66±0,036 |
1,08±0,075 |
0,95±0,066 |
0,90±0,062 |
|
Кишечные |
0,64±0,041 |
0,94±0,071 |
1,16±0,096 |
2,28±0,326 |
2,11±0,273 |
2,03±0,261 |
|
Брыжеечные |
0,66±0,038 |
1,08±0,063 |
1,48±0,166 |
2,21±0,306 |
2,06±0,276 |
1,95±0,244 |
|
Почечные |
0,25±0,005 |
0,64±0,033 |
1,23±0,113 |
1,87±0,230 |
1,14±0,130 |
1,21±0,092 |
|
Печеночные |
0,27±0,006 |
0,67±0,038 |
1,24±0,115 |
2,3±0,332 |
2,09±0,280 |
1,88±0,227 |
|
Паховые |
0,64±0,031 |
1,02±0,079 |
1,95±0,256 |
2,35±0,355 |
2,72±0,493 |
2,18±0,311 |
Таблица №2
Амплитуда сокращений лимфатических узлов у крысят в онтогенезе
|
Лимфатические
узлы |
период
наблюдений, дни |
|||||
|
1 |
5 |
10 |
15 |
20 |
30 |
|
|
Подчелюстные |
0,58±0,040 |
0,79±0,090 |
1,65±0,185 |
2,23±0,323 |
2,19±0,315 |
1,94±0,256 |
|
Подмышечные |
0,58±0,028 |
0,93±0,066 |
1,61±0,174 |
2,19±0,304 |
2,31±0,338 |
2,15±0,299 |
|
Сердечные |
0,2±0,003 |
0,22±0,004 |
0,58±0,033 |
0,98±0,061 |
0,86±0,055 |
0,81±0,051 |
|
Кишечные |
0,54±0,029 |
0,83±0,053 |
1,07±0,081 |
2,18±0,292 |
2,01±0,253 |
1,93±0,223 |
|
Брыжеечные |
0,10±0,002 |
0,65±0,027 |
0,8±0,028 |
2,01±0,205 |
1,86±0,222 |
1,83±0,301 |
|
Почечные |
0,21±0,063 |
0,55±0,025 |
1,13±0,096 |
1,77±0,187 |
1,31±0,110 |
1,11±0,078 |
|
Печеночные |
0,22±0,004 |
0,57±0,038 |
1,09±0,090 |
2,2±0,297 |
1,99±0,251 |
1,78±0,204 |
|
Паховые |
0,54±0,024 |
0,93±0,067 |
1,85±0,239 |
2,14±0,305 |
2,62±0,458 |
1,97±0,264 |
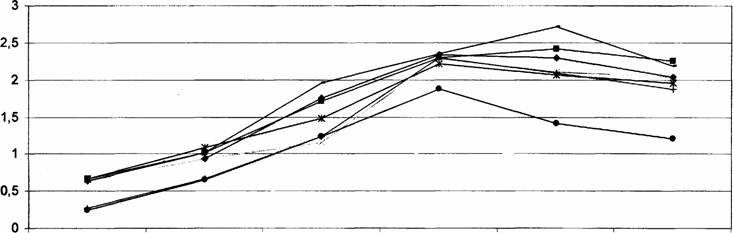
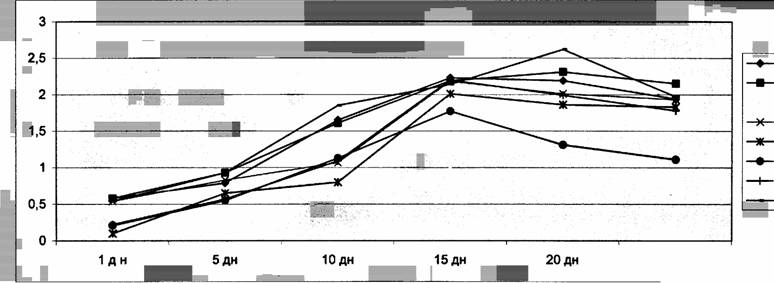
Сказанное хорошо видно на кривой частотной и амплитудной характеристики
лимфатических узлов (рис.1-2).
Как видно из рисунков 1 и
2 направление формирования частотной и
амплитудной характеристик, составляющей спонтанную ритмическую сократительную
активность лимфатических узлов в ходе постнатального развития крыс имеют
аналогичную форму. К моменту рождения
спонтанная сократительная активность всех исследованных лимфатических узлов
низка или практический отсутствует, а наблюдающиеся сокращения нерегулярны и
спонтанны, без определённой закономерности. На 5 день после рождения отмечается
некоторое повышение сократительной деятельности узлов. Однако всё ещё не
обнаруживается регулярность всех компонентов (частотной и амплитудной)
сократительной деятельности лимфатических узлов, хотя общий уровень
биоэлектрической активности заметно повышается.
Кривая амплитуды сокращений
лимфатических узлов у крыс
|
|
|
Подчелюстные Подмышечные Сердечные в Кишечные " Брыжеечные Почечные Печеночные Паховые |
Кривая частоты сокращений лимфатических узлов у крыс
|
|
-Подчелюстные
-Подмышечные -Сердечные -Кишечные -Брыжеечные -Почечные
-Печеночные -Паховые
На 10 день постнатальной жизни
выявляется характерная для органов специфичесность кривой лимфонодуллограммы, сокращения
становятся ритмическими, и выявляется частотно-амплитудная специфичность
сократительной деятельности лимфатических узлов. Начиная с 15 дня и по 30 день обнаруживается
характерное для взрослых крыс картина спонтанной ритмической деятельности
лимфатических узлов. Установлено, что во все
периоды постнатальной жизни частотно-амплитудные характеристики
сократительной деятельности узлов
меняются однонаправлено, т.е. синергично. Наиболее высокая частота и амплитуда сокращении узлов
выявлено на 15 день с последующим сохранением указанных характеристик на 20-30 сутки за исключением почечных лимфатическим
узлов, частотно-амплитудная
характеристика, которого снизилось, в среднем на 40% на 20 и 30 сутки. Следует заметить, что
сократительная деятельность лимфатических узлов такого органа как сердце, обладающего
автоматизмом была низкой по сравнению с сокращением других висцеральных и
соматических узлов. Это, по – видимому связано с тем, что в циркуляций лимфы и
внеклеточной жидкости значительная роль принадлежит мышечному микронасосу сердца
(Косицкий,1975),который работает постоянно.
Таким образом, нами впервые выявлено
становление спонтанной ритмической сократительной активности лимфатических
узлов тела и внутренностей лабораторных крыс, которая имеет ряд особенностей в
отношений различных регионов тела, что в свою очередь позволяет предположить о
регионарной специализации лимфатических узлов (Бикбулатов,1992).