к.б.н. М.В. Горбачева, д.б.н.
Т.Г. Кузнецова
ФГБУН Институт физиологии им. И.П. Павлова РАН,
Санкт-Петербург
Динамика биоэлектрической активности
мозга ребенка 6-7 лет при достижении цели
Аннотация
Установлено,
что стремление к достижению цели, приближающейся с различными скоростями,
активирует поведенческие, вегетативные и центральные реакции саморегуляции. Скорость,
равная 50 мм/с, в отличие от высоких и низких, вызывая реакцию
неопределенности, обладала наибольшей физиологической силой, что проявлялось в
увеличении показателя “стресс-индекса” и наиболее значимой перестройке
биопотенциалов головного мозга. Обнаружено, что усиление активности
α-ритма в ассоциативных зонах правого полушария является общим механизмом
при восприятии эмоционально значимого стимула независимо от возраста человека. В
отличие от взрослых у детей обнаружилась значимая правосторонняя асимметрия
α-ритма в низкочастотном поддиапазоне (7-8 Гц) по всем отведениям, а в
средне- и высокочастотных поддиапазонах (8,75-9,75 и 10,0 до 13,5 Гц соответственно)
– преобладала левосторонняя асимметрия.
Ключевые слова: дошкольники, индекс
напряжения сердечного ритма, когерентность ЭЭГ.
Введение
В 1916 г. на III-м Съезде по
экспериментальной педагогике И.П. Павлов выделил особый рефлекс, назвав его “рефлексом
цели”, подчеркивая, что цели могут быть важными и пустыми и надо отличать
стремление к ее достижению от смысла и ценности цели [9].
Однако до сих пор не ясно, что важнее
физиологическая сила ценности цели или стремление к ней, какова взаимосвязь этих
компонентов в процессе организации целенаправленного поведения, хотя пути ее
решения искали
П.К. Анохин [1], П.В. Симонов [10], H. Heckhausen [13], D. McFarlend [7],
D. Kahneman [3] и др., какие механизмы лежат в основе
целенаправленного поведения, что облегчает, а что затрудняет стремление и
достижение цели, какова роль реакций саморегуляции в организации
целедостижения.
Сложность анализа этой проблемы долгое время
заключалась в отсутствии методик качественной и количественной оценки
обуславливающих ее факторов, что и побудило нас создать методику приближающейся
цели – МПЦ [4], позволившую вскрыть не только содержательную сторону процесса
целедостижения, но и его побудительные и организующие силы, проанализировать
последовательность включения и доминирования активационных (модулирующих)
систем головного мозга – эмоций, ориентировочного рефлекса и сосредоточения,
подойти к анализу реакций саморегуляции, способствующих достижению цели и при
минимальной когнитивной нагрузке моделировать активность этих систем.
Ранее было показано, что скорости
приближения цели-объекта (подкрепления), время ее достижения и расстояние до
нее независимо от вида испытуемого, его пола и возраста активировали
модулирующие системы: эмоций, сосредоточения, ориентировочного рефлекса [11].
Вместе с этим оказалось, что стремление к обладанию (захвату) целью зависит не
только от скорости ее достижения, но и собственной ее ценности [5]. Перечисленные
факторы влияли как на поведенческие, так и на вегетативные реакции
саморегуляции [6].
Встал вопрос, как скорости достижения цели отражаются
на центральном уровне саморегуляции – на изменении биоэлектрической активности
мозга у детей старшего дошкольного возраста.
Была поставлена задача – проанализировать изменения поведенческих, вегетативных и
центральных реакций саморегуляции, и отдельно –асимметричность распределения α-ритма
у детей 6-7 лет при достижении ими цели, приближающейся с различными
скоростями.
Методика
В исследовании с письменного
разрешения родителей принимали участие 6-7-летние дети (n=26) детского сада № 81 г. Санкт-Петербурга.
Использовалась МПЦ [4], основу
которой составляет транспортер длиной 1000 мм, где размещалась достигаемая цель-объект.
Скорость движения цели к испытуемому снижали последовательно: 250-125-50-25-10-5
мм/с с помощью пульта управления, находящегося у исследователя. Перед
испытуемым помещалась кнопка, нажимая на которую, он по команде «Внимание,
работай!» запускал устройство.
В процессе работы велась видеозапись. Анализировались:
длительность реакции сосредоточения, как время активного слежения за движущимся
объектом, рассчитываемое в процентах от нормативного времени достижения цели на
данной скорости, при условии, что коэффициент продуктивности (КП) равнялся
единице, и эмоциональные реакции, меняющиеся в зависимости от скорости движения
объекта.
ЭКГ отводилась от стандартных точек V1, V2.
Анализировался «стресс-индекс» (индекс напряжения – ИН), рассчитываемый по
формуле: ИН = АМо/(2ВР×Мо), где ВР вариационный размах (разница между max
и min значением интервалов R-R), Мо – мода и АМо – амплитуда моды.
Параллельно велась запись ЭЭГ от 10
симметричных отведений правого и левого полушарий: Fp1, Fp2, F3, F4, Р3, Р4,
Т3, Т4, О1, О2, расположенных по
международной схеме 10-20, монополярно с объединенным ушным электродом.
Анализировались корреляционные связи между обозначенными зонами мозга в
исследуемых безартефактных отрезках ЭЭГ продолжительностью не менее 60с и
последующим усреднением по группе испытуемых для каждой из предъявленных
скоростей.
Отдельно рассматривали изменение индекса
α-ритма ЭЭГ для каждой из предъявленных скоростей в диапазоне от 250 до 5
мм/с с последующим усреднением по группе испытуемых и его асимметричность
распределения по отведениям.
Эпоха анализа ЭЭГ составляла 4с, частота
дискретизации 250 Гц. Запись ЭКГ и ЭЭГ вели с помощью 21-канального блока
усилителя биопотенциалов фирмы "Мицар ЭЭГ" (Санкт-Петербург) с портом
для ЭКГ и ЭЭГ-электродов и персонального компьютера. Для анализа результатов
использовали программные пакеты "Mitsar WinEEG" и "Mitsar
WinHRV".
Статистическая обработка осуществлялась в программе StatSoft Statistika 6.0 с применением
непараметрического Т-критерия Вилкоксона, значимыми считались различия на
уровне не менее P<0,05.
Результаты исследования
Достижение
объекта за короткие промежутки времени (4-8с при скоростях 250 и 125 мм/с
соответственно) происходило при спокойном эмоционально положительном
реагировании на ситуацию в целом, не вызывая существенных изменений в ИН
сердечного ритма. Введение скорости приближающегося объекта, равной 50 мм/c, сопровождалось появлением ориентировочной реакции с
негативной окраской и значимым увеличением ИН (p=0,002)
в сравнении с предыдущими состояниями (рис.1, столбики).
При следующих предъявлениях скоростей, приближающегося
объекта, от 25 до 5 мм/с ИН прогрессивно снижался на фоне усиливающегося
эмоционально отрицательного реагирования и появления таких реакций
саморегуляции, как отведение взгляда, отворачивания от установки и т.д. В
результате время слежения (рис. 1, линия) за движущимся объектом со скоростями
10 и 5 мм/с значимо снизилось (p=0,002) относительно
более высоких скоростей приближения объекта (125, 50 и 25 мм/с).
Анализ
ЭЭГ показал, что распределение значимых (r>0,4-0,8)
корреляций как в спокойном состоянии при открытых глазах (ГО), так и в ответ на
скорости движения объекта, равные 250, 125, 25, 10 и 5 мм/с, оказалось сходным
(рис.1, топограммы). Наиболее часто взаимосвязанными оказывались симметричные
лобно-моторные (17%–23%), левосторонние теменно-затылочные (от 16%–24%) и правосторонние
височно-теменно-затылочные (от 16%–26%) зоны мозга, при этом установились
межполушарные лобно-теменные связи (от 19%–28%).
** ** * *
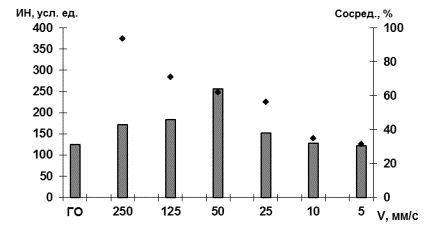
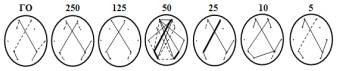
Рис.1. Изменение ИН,
длительности реакций сосредоточения внимания на движущемся объекте,
распределение корреляционных связей при снижении скорости ее приближения.
Обозначения:
левая ось ординат – ИН в усл. ед.; правая ось – длительность реакций слежения в
%; столбики – ИН усл. ед.; линия – изменение активного времени слежения
(сосредоточения внимания) за объектом; ось абсцисс – скорости движения объекта,
мм/с. Под графиком – топограммы распределения корреляционных связей между
исследуемыми зонами мозга. Пунктирные линии – от15% до 19% связей, черные
тонкие – от 20% до 29% связей, черные жирные – больше 30% связей. * –
p<0,05; ** – p<0,01.
Существенные изменения в распределении
корреляционных связей наблюдались при достижении объекта, приближающегося со
скоростью, равной 50 мм/с, когда практически все исследуемые зоны мозга
оказались взаимосвязанными на фоне увеличения количества связей между ними. По
сравнению с распределением значимых связей, зарегистрированных в ответ на
введение высоких и низких скоростей, сохранилось и/или увеличилось количество
правосторонних височно-теменно-затылочных (от 30%–40%) и левосторонних
теменно-затылочных связей (45%), удвоилось число межполушарных лобно-теменных
(с 20 до 40%), появились новые межполушарные лобно-лобные, лобно-затылочные (до
25%) и лобно-височные (25%) связи.
Таким образом, в работе еще раз доказано,
что поведенческие реакции саморегуляции (пассивное избегание), возвращая
организм в спокойное (положительное) гомеостатическое состояние, способствуют
нормализации как вегетативных (снижение стресс-индекса), так и ЭЭГ (снижение
внутри- и межполушарной активации биопотенциалов, но без выраженной
асимметричности их распределения) ответных реакций при возникающих затруднениях
в процессе стремления к достижению цели.
В то же время неопределённость ситуации
(внезапное снижение скорости до 50 мм/с), на фоне выраженной реакции
беспокойства и негативно-ориентировочной реакции («Что случилось? Почему?»), привела
к достоверному возрастанию показателя «стресс-индекса» механизмов регуляции сердечного
ритма, значимой перестройке биопотенциалов головного мозга и усилению
корреляции между отдельными его зонами во всех частотных диапазонах.
А вот анализ α-ритма выявил иную динамику. В целом изменение
индекса α-ритма в полном диапазоне (8-13 Гц) существенно не менялась, что
согласуется с данными иностранных авторов [14], полученными на взрослых людях при
"пассивном" просмотре IAPS стимулов (International Affective Picture System, University of Florida, 1999).
У детей же в нашем исследовании при
восприятии эмоциогенных стимулов – достижении цели при различных скоростях ее
движения – асимметричность α-ритма проявилась по поддиапазонам.
Известно, что этот ритм у человека
окончательно формируется к 15-17 годам. Однако уже в 7-летнем возрасте появляется
низкочастотный α-ритм (не более 8 Гц), но без «веретенообразных» модуляций
и градиента падения от затылочных зон к лобным. Ряд детских нейрофизиологов [2,
8, 12] этот ритм подразделяет на низкочастотный (7,0 – 8,5 Гц), среднечастотный
(8,75 – 9,75Гц) и высокочастотный (10,0 – 13,5 Гц).
Так, у 77% 6-7-летних детей с
низкочастотным диапазоном α-ритма наблюдалась сходная с фоном, значимая
правосторонняя асимметрия практически по всем отведениям вне зависимости от
активации модулирующих систем, определяемых скоростью достижения объекта (табл.1,
столбцы 1, 4, 7 и 10).
Таблица 1.
|
Номера столбцов |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
|
Диапазоны
частот α-ритма, Гц |
7,0-8,5 |
8,75-9,75 |
10,0-13,5 |
7,0-8,5 |
8,75-9,75 |
10,0-13,5 |
7,0-8,5 |
8,75-9,75 |
10,0-13,5 |
7,0-8,5 |
8,75-9,75 |
10,0-13,5 |
|
Топограммы |
|
|
|
|
||||||||
|
Состояния |
Фон (ГО) |
250-125 мм/с |
50-25 мм/с |
10-5 мм/с |
||||||||
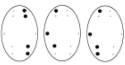
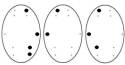
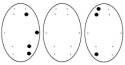
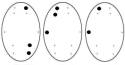
Обозначения: топограммы распределения α-ритма ЭЭГ по изучаемым отведениям
у детей 6-7 лет в процессе достижения им цели, соответствующие: Фон – исходное состояние (ГО). Цифры под топограммами – скорости
достижение цели: 250-125 мм/с (эмоционально
положительное состояние); 50-25 мм/с (состояние
неопределенности); 10-5 мм/с (эмоционально отрицательное состояние).
Точки на топограммах – отведения, в
которых отмечалась достоверная (p<0,02) частотная асимметричность
α-ритма ЭЭГ в Гц.
У 11-12% детей со средне- (табл.1,
столбцы 2, 5, 8 и 11) и высокочастотным (табл.1, столбцы 3, 6, 9 и 12)
α-ритмом появилась тенденция к установлению левосторонней асимметрии
преимущественно в лобных, височных и теменных отведениях в исходном состоянии и
не зависимо от эмоциональной окраски, определяемой скоростью движения объекта.
Таким образом, в работе показана важная роль
поведенческих реакций саморегуляции в нормализации функционального состояния
ребенка, приводя «стресс-индекс» механизмов регуляции сердечного ритма и ЭЭГ к
исходному состоянию.
Существенные перестройки общей биоэлектрической
активности и усиление активности α-ритма в ассоциативных зонах правого
полушария и асимметричное его распределения в теменно-височных областях у
испытуемых 6-7 лет, равно как и у взрослых людей, сопряжены с возникновением
негативно-ориентировочной реакции. Выявленная же асимметричность по
поддиапазонам α-ритма у детей, скорее является показателем зрелости коры
головного мозга, чем реакцией на эмоциогенность стимула.
Литература
1.
Анохин
П.К. Рефлекс цели как объект физиологического анализа // Журн. высш. нерв. деят. – 1962. – Т.12. – Вып.1. – С. 7-21.
2.
Безркуких
М.М., Мачинская Р.И., Фарбер Д.А. Структурно-функциональная
организация развивающегося мозга и формирование познавательной деятельности в
онтогенезе ребенка // Физиология человека. – 2009. – Т. 35. – №6. – С.
10-24.
3.
Канеман
Д. (Kahneman D.) Внимание и
усилие. – М.: Смысл, 2006. – 287 с.
4.
Кузнецова
Т.Г., Сыренский В.И., Наулайнен Б.А. Способ определения целеустремленности:
А.с. 1410948 СССР. МКИ4 А61В5/16/ // Открытия. Изобретения. – 1988.
– № 27. – С. 44-46.
5.
Кузнецова
Т.Г., Горбачева М.В. Сравнительный анализ некоторых поведенческих реакций у
детей и шимпанзе в процессе достижения ими цели с различной субъективной
ценностью // Вестник Самарского госуниверситета. Естественнонаучная серия. –
2012. – №3/1 (94). – С. 192-202.
6.
Кузнецова
Т.Г., Горбачева М.В. Роль реакций саморегуляции в организации целенаправленного
поведения детей 6-7 лет // Международный научно-исследовательский журнал. Часть
1. – 2013. – № 12 (19). – С. 23-28.
7.
Мак-Фарленд
Д. (McFarlend D.)
Поведение животных. – М.: Мир, 1988. – 487 с.
8.
Мачинская
Р.И., Соколова Е.С., Крупская Е.В. Формирование функциональной организации коры
больших полушарий в покое у детей младшего школьного возраста с различной
степенью зрелости регуляторных систем мозга. Сообщ. 2. Анализ когерентности
альфа-ритма // Физиология человека. – 2007. – Т.33. – №2. – С. 5-15.
9.
Павлов
И.П. Рефлекс цели / Рефлекс свободы. – СПб: Питер, 2001. – С. 68-75.
10.
Симонов
П.В. Мотивированный мозг. – М.: Наука, 1987. – 270 с.
11.
Сыренский
В.И., Кузнецова Т.Г. Рефлекс цели у приматов. – Л.: Наука, 1990. – 120 c.
12.
Фарбер Д.А., Бетелева Т.Г., Горев А.С. и др. Функциональная
организация развивающегося мозга и формирование когнитивной деятельности //
Физиология развития ребенка: теоретические и прикладные аспекты. М.: НПО
«Образование от А до Я», 2000. – С. 82-103.
13.
Хекхаузен
Х. (Heckhausen H.)
Мотивация и деятельность: пер. с нем. – Т.1. М.: Педагогика, 1986. – 408 с.
14.
Muller
M.M., Keil A., Gruber Т., Elbert T. Processing of
affective pictures modulates right-hemispheric gamma band EEG activity // Clin.
Neurophysiol. – 1999. – V. 110. – P. 1913-1920.