Биологические
науки/8. Физиология человека и животных.
Институт физиологически активных
веществ РАН, Россия
Кохан
В. С., Болкунов А. В., Иванова Н.А., Ванькин Г. И.
Некоторые особенности
поведения мышей – нокаутов по гену
гамма-синуклеина
Гамма(γ)-синуклеин
– представитель семейства синуклеинов,
цитоплазматических, преимущественно нейрональных белков, которые в эволюции
появились поздно и имеются только у позвоночных. Значительные количества γ-синуклеина обнаруживаются в аксонах и
пресинаптических окончаниях нейронов областей, ответственных за формирование
эмоций и памяти. Роль синуклеинов в
функционировании нервной системы до сих пор не выяснена, но установлено, что
нарушение метаболизма этих белков ведет к развитию различного типа
патологий [1, 2].
Семейство синуклеинов включает три белка (альфа(α)-,
бета(β)- и гамма(γ)-синуклеин) со сходным планом строения молекулы и
почти идентичными аминокислотными последовательностями N-концевых областей.
Синуклеины кодируются тремя различными генами, имеющими высокую степень
гомологии [3].
Для
изучения функции
синуклеинов большую ценность представляют линии мышей, полученные путем
направленной модификации их генома, приводящей, к инактивации одного из
синуклеинов (нокаутные животные). Получены α-, β- и γ-синуклеин
нокаутные мыши [1]. Нокауты по γ-синуклеину (γ-нокауты)
жизнеспособны, фертильны и не имеют выраженного фенотипа, выявляемого без
применения инструментальных методов анализа. По последним данным фенотипу γ-нокаутов присущи сниженный уровень
ситуативной тревожности, повышенные исследовательская активность и склонность к
риску в сравнении с мышами, имеющими интактный ген [4].
Целью
настоящей работы было исследовать такие
особенности поведения животных, как
поколачивание хвостом (tail rattling), зависание (floating) в водном лабиринте Морриса, а также акты дефекации и
уринации в условиях новой обстановки.
При
осуществлении различных поведенческих тестов зачастую фиксируются лишь
избранные акты, исходя из цели исследования. В процессе детекции таких
параметров учитывается лишь факт наступления того или иного события, но не характер его
исполнения или действия его
сопровождающие. Несомненно, для наблюдения многих особенностей поведения
существуют специализированные методики, однако первичную картину таких поведенческих
актов можно получить и при детальном анализе неспецифических тестов. В трёх
тестах, а именно в тесте выработки условного рефлекса избегания (челночная
камера), узнавания нового объекта, водном лабиринте Морриса, нами и были
отмечены особенности в поведении
γ-нокаутов.
Получение
мышей – γ-синуклеиновых нокаутов (γ-нокаутов), описаны ранее [1]. В данной
работе были использованы мыши на генетической основе линии С57Bl-6J.
Во всех
экспериментах были использованы самцы в возрасте 6 месяцев массой 22-24 г,
которых содержали по одному в клетке на протяжении всей программы
исследований при температуре 220С в условиях естественного 12
часового светового цикла, в свободном доступе к пище и воде. Генетический
анализ экспериментальных животных проводили, как описано ренее [1]. Группы
формировали по 10 животных.
Статистическую
обработку данных производили с помощью программы "Statistica 6.0". Достоверность различий выборок определяли с помощью t-критерия Стьюдента для независимых переменных.
Работы
с животными проводили в соответствии с "Правилами лабораторной практики в Российской
Федерации" от 2003 года.
При
проведении теста выработки условного рефлекса активного избегания в челночной
камере была отмечена такая особенность поведения, как поколачивание хвостом –
с третьего и всех последующих этапов обучения (всего восемь). Тест
выполнялся по стандартной методике, перерыв между повторами составлял 24 часа
[5]. Полученные данные суммировали и усредняли.
Акты
дефекации и уринации были зафиксированы на первом этапе теста узнавание новой
локации объекта – в процессе 15 минутного ознакомления с камерой. Размер
камеры составлял 48х38х30 см. Тест был выполнен по стандартной методике [6].
Полученные данные усредняли в пересчёте на одну мышь.
Зависание
в водном лабиринте Морриса представляло собой замирание мыши и удерживание на
воде в свободном дрейфе более 5 с. Тест выполнялся по стандартной методике в
бассейне диаметром 80 см, освещённость составляла 25 лк [7]. Зависания
фиксировали на всех этапах обучения, данные усредняли.
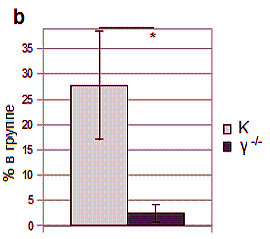
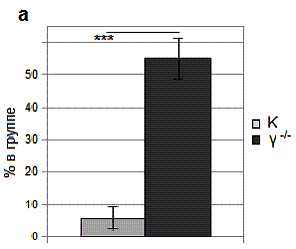
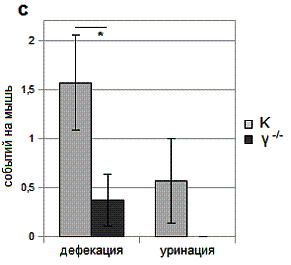
Рис. 1. Особенности поведения γ-нокаутных мышей.
a – поколачивание хвостом
в челночной камере.
b – зависание в водном
лабиринте Морриса.
c – акты уринации и
дефекации в условиях новизны.
К –
контрольная группа животных, содержащих нормальный немодифицированный ген
γ-синуклеина, γ-/- – нокауты по гену γ-синуклеина.
* – p<0,05, ** – p<0,01, *** – p<0,0001.
Данные о поколачивании хвостом представлены на рис. 1a.
Только 5.7 % мышей контрольной группы осуществляли акты поколачивания хвостом,
тогда как в группе γ-нокаутов такое поведение
было свойственно 55 % животных, что на
49.3 % (p = 0.00014) больше
контроля. Следует отметить, что поколачивание хвостом животными контрольной
группы было хаотичным, тогда как γ-нокауты поколачивали либо непосредственно
до сигнала к переходу, либо осуществив переход в нормативное время. При
получении удара током и в момент перерыва между повторами поколачивание хвостом
не отмечалось.
Зависание в водном лабиринте Морриса отображено
на рис. 1b. Животные контрольной группы
зависали в бассейне на 25.9 % (p=0.0267) чаще, чем
группа γ-нокаутных мышей. Какая либо динамика зависания контрольных мышей
в повторах обучения не отмечена, тогда как в группе гамма γ-нокаутов
зависания были только при первом подходе.
Данные об актах дефекации и уринации
представлены на рис. 1c. Активность
дефекации γ-нокаутов составляла в 4.1 (p=0.04) раза меньше в сравнении с контролем. Активность уринации
достоверно не различалась, однако следует отметить её полное отсутствие в
группе мышей γ-нокаутов в течение
всех 15 минут наблюдения.
Известно, что акты дефекации и уринации
непосредственно связаны с уровнем тревожности, необходимо также учесть, что их
детекция проводилась нами на первом этапе теста, а следовательно — в условиях
вынужденной новизны. Таким образом, мыши, нокауты по гену γ-синуклеина обладают сниженной
ситуативной тревожностью по сравнению с контрольной группой животных.
Исходя из того, что γ-нокауты осуществляли акты поколачивания хвостом в
приближении или после избегания угрозы (удар током), мы склонны думать, что
такое поведение говорит как о сниженном уровне страха (базовой тревожности),
так и о склонности к агрессии (что требует дополнительных исследований) по
сравнению с контролем.
Практически полное отсутствие зависания в водном
лабиринте Морриса γ-нокаутов
свидетельствует, на наш взгляд, о высоком пороге входа в депрессивное состояние
по сравнению с контролем [8].
Таким образом отсутствие гена γ-синуклеина у мышей приводит к
снижению ситуативной и базовой тревожности, снижению уровня страха и повышению
порога депрессивного статуса. Что находит подтверждение в наших предыдущих
исследованиях.
Литература:
1.
Ninkina N. N., Papachroni, K., et
al. Neurons expressing the highest levels of gamma-synuclein are unaffected by
targeted inactivation of the gene. Mol. Cell. Biol.. 2003. 23: 8233–824.
2. Dev K. K., Hofele K., Barbieri S., Buchman V. L., van der Putten H. Part II: alpha-synuclein and its molecular pathophysiological role in
neurodegenerative disease. Neuropharmacology. 2003. 45(1): 14-44.
3. Нинкина Н. Н., Бухман В. Л., Синуклеины - иметь или не иметь? Генетика. 2000. Том 36, № 11, стр. 1487-1492.
4. Кохан В. С., Болкунов А. В. et al. Исследование эмоционально
опосредованного поведения мышей – нокаутов по гену гамма-синуклеина. Журн.
высш. нерв. деят. 2010 [в печати].
5. Буреш Я., Бурешова О., Хьюстон П. Методики и основные
эксперименты по изучению мозга и поведения. Москва. «Высшая школа». 1991. 399 с.
6. Ennaceur A,
Delacour J. A new one-trial test for neurobiological studies of memory in rats.
I. Behavioral data. Behav Brain Res. 1988. 31: 47–59.
7. Morris RGM.
1981. Spatial localization does not reguire the presence local cues. Learning
and motivation. 12: 239 – 250.
8. Strekalova
T., Spanagel R., et al. Stress-induced anhedonia in mice is associated with
deficits in forced swimming and exploration. Neuropsychopharmacology. 2004
Nov;29(11):2007-17.