ЭПИТЕЛИАЛЬНО-СТРОМАЛЬНЫЕ
МОРФОФУНКЦИОНАЛЬНЫЕ ЗОНЫ ОПРЕДЕЛЯЮТ ПРОЛИФЕРАЦИЮ И ДИФФЕРЕНЦИРОВКУ КАМБИАЛЬНЫХ
КЛЕТОК
Явишева
Т.М., Щербаков С.Д.
ЗАО
«Р-ФАРМ», лаборатория механизмов регуляции стволовых клеток
В данном работе обобщен собственный
материал, полученный при изучении особенностей пространственной организации
разных эпителиев (переднего и заднего эпителия роговицы глаза, эпидермиса
кожи), стромальной ткани у животных и человека в возрастном аспекте, опухолевой
ткани у больных раком легкого, пищевода, молочной железы.
Пролиферация и дифференцировка камбиальных
клеток тесно связана с работой морфофункциональных зон. Действительно,
камбиальные клетки функционируют не
хаотично, а в специальных зонах,
обеспечивающих их жизнедеятельность. Морфофункциональная зона включает два
компонента – эпителиальный и стромальный, которые оказывают друг на друга
противоположный эффект за счет ключевых белков
RhoA (один из малых G-белков) и Src-тирозинкиназы:
эпителиальные клетки вызывают спастическое действие, так как в них преобладает
экпрессия белка RhoA над Src , а стромальные –
релаксирующее, потому что в них,
наоборот, преобладает экспрессия Src над RhoA (рис.1).
Каждый компонент зоны в свою очередь состоит из двух субъединиц, которые работают поочередно и включают по 12 камбиальных клеток.
При делении 12 камбиальных клеток образуются
пары материнских и дочерних клеток, создающих электрическое поле, в котором
происходит растяжение дочерних клеток и их готовность к дифференцировке, так
как при этом выпетливаются определенные участки хромосом. 6 пар материнских и
дочерних клеток создают электрическое поле пороговой величины, при котором
растяжения и дифференцировки дочерних клеток не происходит. Растяжение дочерних
клеток в электрическом поле приводит к возбуждению в них SH2
домена Src , который участвует в построении их цитоскелета –
микротрубочек и промежуточных филаментов, необходимых для дальнейшей
дифференцировки.
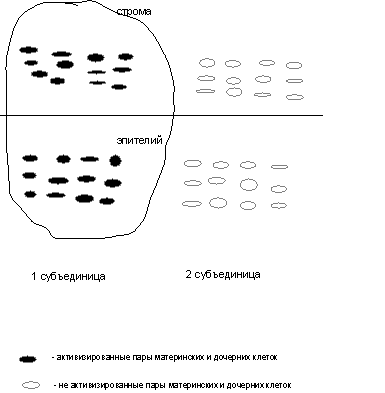
Рис. 1.
Схематическое изображение работы морфофункциональной зоны в нормальной
эпителиальной ткани. Взаимодействие стромального и эпителиального
компонентов одной из субъединиц зоны.
Следовательно,
функция камбиальных клеток в морфофункциональной зоне сводится в первую очередь
к экспрессии в дочерних клетках этого домена. У каждого индивидуума имеется
свой конституциональный уровень экспрессии Src , который закладывается
в эмбриогенезе и колеблется в пределах нормы. Поэтому камбиальные клетки определенного индивидуума поддерживают такой
уровень экспрессии Src в организме, который свойственен только данному человеку.
Таким образом, численность камбиальных клеток в морфофункциональной зоне должна строго
контролироваться. Действительно, если число их будет не больше 6, то дочерние
клетки не будут подвергаться дифференцировке, что может привести к развитию
злокачественной опухоли. Контроль численности камбиальных клеток вытекает из самих принципов работы морфофункциональной
зоны. Камбиальные клетки возникли из презумптивной эктодермы на границе сильно
экспрессированной RhoA и практически отсутствующей экпрессии Src,
поэтому отсутствует какая-либо дифференцировка этих клеток (рис. 3 в). В связи
с тем, что в камбиальных клетках не экспрессирован SH2 домен Src,
участвующий в образовании цитоскелета клетки, то в них не образуется
микрофиламентов, позволяющих им распластываться на базальной мембране, поэтому
камбиальные клетки имеют вертикальную по отношению к базальной мембране ось
деления. Отсутствие в камбиальных
клетках экспрессии SH2
домена Src, участвующего в
образовании микротрубочек веретена деления, приводит к тому, что в камбиальных
клетках равномерное распределение хромосомного материала между материнской и
дочерней клеткой, получившихся при делении, происходит за счет тянущей силы
электрического поля, возникающего между хромосомами ядер этих клеток. Дочерняя клетка до растяжения в электрическом поле ничем не отличается от
материнской. При делении первых 6 камбиальных клеток дочерние клетки не
подвергаются растяжению, поэтому каждая из них может стать камбиальной и быть
источником роста. Однако при делении
следующих 6 камбиальных клеток образуется уже 12 пар материнских и дочерних
клеток, которые создают необходимое электрическое поле, растягивающее все 12
дочерних клеток. Таким образом, в морфофункциональной зоне не может
образоваться ни одна лишняя камбиальная клетка. В случае снижения численности
камбиальных клеток в морфофункциональной зоне по тем или иным причинам,
камбиальные клетки будут делиться до тех пор, пока численность их ни станет 12 , что приведет к остановке дальнейшего увеличения их количества
за счет начала дифференцировки дочерних клеток в электрическом поле.
Пролиферация же камбиальных клеток и их
потомков в морфофункциональной зоне начинается лишь тогда, когда уровень экспрессии
RhoA в этих клетках будет выше
умеренного преобладать над Src (рис. 2) .
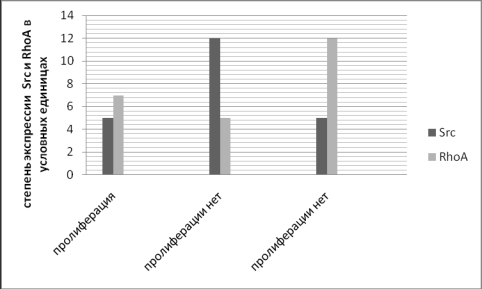
Рис. 2. Схематическое
изображение экспрессии Src-киназы и RhoA
по отношению друг к другу, обусловливающих условия проиферации клеток.
Это
связано с тем, что Src
инактивирует часть белка RhoA , оставшаяся же часть
этого белка будет участвовать в развитии сократительного аппарата клетки,
необходимого для цитокинеза. Такое условие создается при функционировании обоих компонентов морфофункциональной зоны – эпителиального и стромального. Так, стромальные
дочерние клетки первыми уходят из зоны, так как их главная ось в отличие от
эпителиальных клеток, ориентирована вглубь ткани, поэтому в зоне начинают преобладать эпителиальные
дочерние клетки, главная ось которых параллельна базальной мембране. За счет
превалирования последних в
морфофункциональной зоне экспрессия RhoA будет выше умеренного преобладать над Src,
что обусловит пролиферацию как камбиальных, так и остальных клеток
морфофункциональной зоны. При других условиях пролиферации не будет. Так,
если в зоне чрезмерно усилится
активность Src-киназы, то это приведет
к значительной инактивации белка RhoA, что вызовет ослабление
пролиферации. Если экспрессия RhoA
будет существенно выше активности Src, то усилится спазм клеток вплоть до полной остановки пролиферации.
Следовательно, работа морфофункциональных
зон по пролиферации и дифференцировки
камбиальных клеток является автономной.
При этом электрическое поле, образующееся при делении камбиальных клеток, подготавливает
дочерние клетки к дифференцировке, т.е. возбуждает в них SH2
домен Src , необходимый для построения цитоскелета, а микроокружение камбиальных элементов в
морфофункциональной зоне за счет
вырабатываемых им биологически активных факторов, в которых имеется
определенное соотношения белков Src
и RhoA , производит дальнейшую
тонкую дифференцировку клетки, соответствующую определенному виду ткани. Таким
образом, клетки морфофункциональной зоны, окружающие камбиальные клетки,
определяют функцию последних. На примере эмбриогенеза видно, что клетки
эпидермиса возникли из эктодермы путем принудительного растягивания их в
поперечном направлении во время движения
нервных валиков навстречу друг другу, при этом главная ось клеток эктодермы из вертикального положения
переходит в горизонтальное, что приводит к раскручиванию уже других локусов хромосом, обеспечивая
эпидермальную дифференцировку (рис. 3 б).
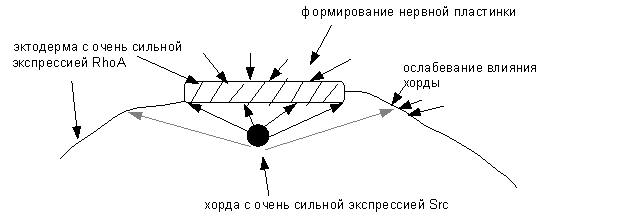
а
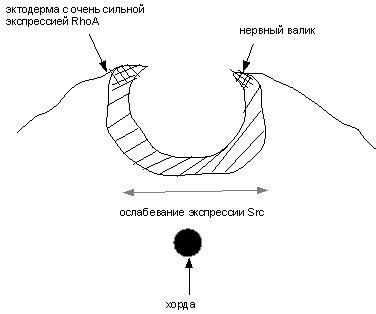
б
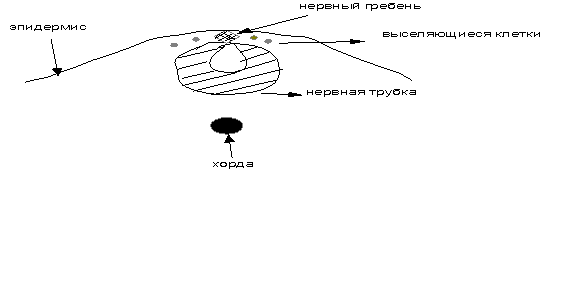
в
Рис. 3 (а-в). Схема образования нервной трубки,
нервного гребня и эпидермиса
а – формирование нервной пластинки ; б - образование нервных валиков и изгибание
нервной пластинки; в – формирование нервной трубки, эпидермиса и нервного гребня, из которого выселяются
камбиальные клетки
Следовательно,
в процессе эмбриогенеза происходит моделировка тканей, при этом в каждой ткани
устанавливается свои, только данной ткани присущие взаимоотношения ключевых
белков – Src и RhoA; впоследствии же
камбиальные клетки, возникшие из эктодермы на границе сильно экспрессированного
RhoA и слабо выраженной Src -киназы, выселяются в
различные ткани, обеспечивая их регенерацию и базовый уровень экспрессии SH2
домена Src-киназы в их клетках
и являются полипотентпыми (рис. 3 в).
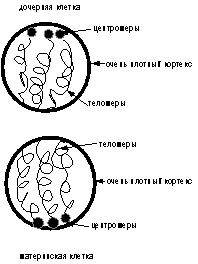
а
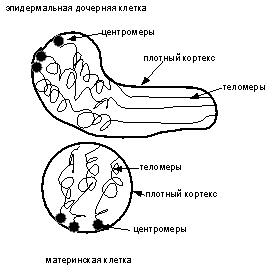
б
Рис.
4 (а, б). Схематическое изображение выхода камбиальной клетки из митоза (а) и
образования эпидермальной дочерней клетки (б).
Так, если камбиальные клетки
функционируют в окружении эпителиального компонента зоны, например,
эпидермиса, то базовый уровень SH2 домена в дочерних
клетках, получившихся при делении камбиальных клеток, будет корректироваться соответствующим микроокружением. Это приведет
к тому, что экспрессия белка RhoA
в дочерних клетках будет умеренно
преобладать над экспрессией Src, как и в окружающих их
эпидермальных клетках. Поэтому образовавшиеся в электрическом поле
микротрубочки и промежуточные филаменты
дочерней клетки растягивают нижний полюс ядра этой клетки, выпетливая
хромосомы ближе к теломерам, принципиально определяя таким образом эпителиальную дифференцировку (рис. 4 а, б). Для
того чтобы эпителиальные клетки вступили в пролиферацию необходимо лишь
незначительно повысить уровень RhoA в них, чтобы
RhoA выше умеренного превышал уровень Src.
Поэтому эпителиальные клетки обладают
высокой пролиферативной активностью.
Если же камбиальные клетки находятся в окружении стромальных клеток, в которых экспрессия Src умеренно превышает RhoA , то в дочерних клетках,
получившихся при делении камбиальных, за счет микроокружения будет
индуцироваться такая же экспрессия этих белков. Это приведет к релаксации
кортекса стромальных дочерних клеток (рис. 5 а, б) и усилению растяжения их
силами электрического поля, что обусловит выпетливание хромосом ближе к
центромерам, определяя мезодермальную дифференцировку. Для вступления этих
клеток в митоз необходимо уже значительное повышение уровня экспрессии в них RhoA таким образом, чтобы этот белок выше умеренного преобладал над Src в этих клетках. Это обусловливает более низкую пролиферативную активность
фибробластов по сравнению с эпителиальными клетками.
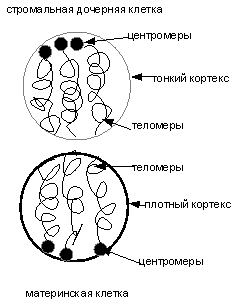
а
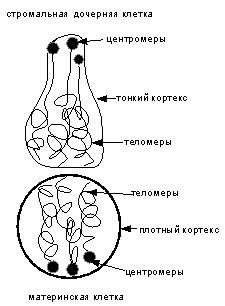
б
Рис.
5 (а, б). Схематическое изображение стромальной дочерней клетки (а) и
растяжения ее в электрическом поле (б).
В нервных клетках экспрессия RhoA резко преобладает над Src,
что приводит к спазму клеток и блоку пролиферации. Это связано с тем, что в
период
эмбриогенеза в нервной пластинке, образующейся из эктодермы, под
влиянием на нее клеток хорды, происходит быстрая, но транзиторная экспрессия Src-киназы
(рис. 3 а). Для того, чтобы нервные клетки начали пролиферировать, необходимо чтобы
RhoA выше
умеренного преобладал над Src, а для этого необходимо
повысить уровень Src-киназы в них.
Несмотря на автономность работы, с
возрастом происходит снижение численности камбиальных клеток в
морфофункциональной зоне. Это связано с функционированием важнейшей
морфофункциональной зоны – гипоталамуса. С наступлением пубертатного периода
постепенно усиливается влияние
эстрогенов на головной мозг, которые активизируют Src-киназу, что снижает
RhoA активность головного мозга. В результате гормональная активность
половых желез сначала увеличивается и наступает гиперэстрогенемия, а затем истощается, что приводит к гипоэстрогенемии.
В период гиперэстрогенемии усиливается влияние Src–киназы на клетки
периферических органов. Так, экспрессия Src -киназы в еще большей
степени усилится в фибробластах в этот период, а в эпителиальных клетках – снизится
RhoA активность, так как Src в некоторой степени инактивирует RhoA.
Следовательно, в этот период усиливается влияние стромального компонента зоны
на эпителиальный. Однако усиление
экспрессии Src в
фибробластах снизит их пролиферативную активность, так как для вступления этих
клеток в митоз необходимо значительное повышение RhoA в этих клетках. Пролиферативная активность эпителиальных
клеток также страдает при гиперэстрогенемии, так как сниженная экспрессия RhoA
в них требует теперь большей
активизации этого белка при
вступлении этих клеток в митоз. Поэтому
в период гиперэстрогенемии в связи с ухудшением пролиферации эпителиальных и стромальных клеток при
действии различных неблагоприятных условий создаются условия для развития
сарком и раковой опухоли.
Для гипоэстрогенемии характерно снижение
влияния стромы на эпителий, так как в фибробластах падает экспрессия Src.
При этом фибробласты пожилых людей способны быстрее вступать в митоз, чем молодых,
потому что для пролиферации
фибробластов теперь требуется в меньшей степени экспрессировать RhoA
. В
связи с этим у пожилых пациентов качество рубца лучше, чем у молодых. Снижение
влияния стромы на эпителиальную ткань приводит к значительному усилению RhoA
в клетках эпителия. Нарастает спазм клеток, что уменьшает их пролиферативную
активность. Ригидность кортикальных слоев, мешающая пролиферации и
дифференцировке, не может быть устранена слабым влиянием стромы в этот период.
Следовательно, эпителиальный компонент морфофункциональной зоны страдает в
большей степени, чем стромальный. Именно поэтому, первое сокращение количества
камбиальных клеток до 11 в эпителиальном компоненте морфофункциональной зоны происходит в период гипоэстрогенемии,
т.е. после 60 лет. У людей старше 75
лет при нарастании ригидности клеток эпителиальной морфофункциональной зоны
численность камбиальных клеток падает
до 7 и приближается к пороговому
уровню (6 клеток), при котором дифференцировка отсутствует. В связи с этим у
пожилых людей злокачественные опухоли встречаются чаще, чем у молодых, причем,
эпителиальные опухоли превалируют над стромальными, так как эпителиальный
компонент морфофункциональной зоны страдает в большей степени. Однако у пожилых
людей в связи с ослаблением влияния стромы на эпителий и ригидностью клеток, в
опухолевой ткани пролиферативные процессы также будут снижены, что приводит к
более торпидному течению опухолевого процесса.
Таким
образом, процессы пролиферации и дифференцировки клеток являются
взаимосвязанными и осуществляются в морфофункциональных зонах, причем, окружающая ткань обусловливает и конкретную
дифференцировку, и пролиферацию камбиальных клеток: дифференцировку - за счет
возбуждения в
дочерних клетках соответствующей
этой ткани эспрессии белков Src
и RhoA, а пролиферацию - за
счет повышения в клетках зоны экспрессии RhoA выше умеренного по
сравнению с Src.