К.б.н. Атамбаева Ш.А., д.б.н. Иващенко
А.Т., д.б.н. Карпенюк Т.А.
к.б.н. Гончарова А.В., Хайленко В.А.
НИИ проблем биологии и биотехнологии Казахского национального
университета имени аль-Фараби, Казахстан
Свойства интронов и
экзонов в генах из различных участков хромосом Сaenorhabditis elegans
В настоящее время
проблема выяснения функциональной роли интронов и их свойств в различных генах
и геномах активно разрабатывается. В генах эукариотических клеток экзоны и
интроны имеют много особенностей, которые способствуют выяснению биологической
функции интронов [1-3]. Соотношение длин интронов и экзонов, а также число
интронов в гене могут существенно влиять на время сплайсинга пре-мРНК и,
соответственно, на экспрессию генов [4]. Представляется важным выяснить,
наблюдается ли изменение свойств экзонов и интронов в зависимости от
хромосомной локализации и числа интронов в генах ядерного генома C. elegans.
Нуклеотидные
последовательности хромосом I-V и Х хромосом ядерного генома C. elegans были получены из базы данных (http://www.wormbase.org). Из каждой хромосомы формировали
группы генов содержащих 1-5 интронов, а также группы генов с 6-9 и с 15 и более
интронами. Анализ частоты встречаемости длины интронов и экзонов проводили в
интервалах 1 – 20 н., 21 – 40 н., 41 - 60 н. и так далее до 400 н., а также
более 400 н. Для анализа свойств генов в зависимости от плотности генов были
выбраны четыре участка с низкой плотностью генов (УНПГ). Наиболее высокой средней плотностью генов на участок обладает хромосома
II – 75 генов, а
наиболее низкой плотностью хромосома Х – 49 генов.
В генах C. elegans, содержащих различное число
интронов, средняя длина экзонов в генах I-V хромосом независимо от числа интронов в генах была близкой:
для хромосом I и II - 241±21 н., III -
227±26 н., IV - 228±33 н. и V - 237±33 н. В генах хромосомы Х средняя длина
экзонов была меньше, чем в остальных хромосомах - 194±21 н. Средняя длина
интронов в генах не зависела от числа интронов в гене. Так, в хромосомах I и II она равнялась 386±60 н., в III, IV и Х
хромосомах она составляла соответственно 338±38 н., 336±53 н. и 251±49 н. В
хромосоме V средняя длина интронов в
генах была ниже, чем в других хромосомах - 201±13 н. Таким образом, средние
длины интронов и экзонов в генах C. elegans не зависели от числа интронов в генах каждой хромосомы, но отличались для
генов из некоторых хромосом. Сумма экзонов в генах с одним интроном варьировала
от 469 н. (хромосома Х) до 579 н. (хромосома V). По мере увеличения числа интронов в генах
сумма длин экзонов в них возрастала в несколько раз и равнялась для генов
указанных выше хромосом соответственно 3908 и 5693 н. Общая длина гена в каждой
из хромосом генома C. elegans увеличивалась в 12,1-15,8 раз с увеличением
количества интронов в гене. Доля экзонов и интронов, различающихся по длине в
интервалах 1 - 400 н. и более 400 н., во всех группах генов изменялась
специфично. В генах всех хромосом C. elegans доля экзонов с длиной более 400 н. уменьшалась с увеличением числа
интронов в гене до 5. При этом доля экзонов с длиной в интервале 60-180 н.
одновременно увеличивалась (рис.1).
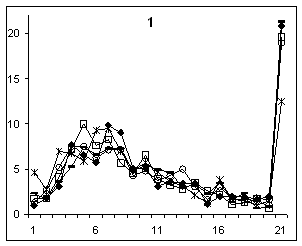
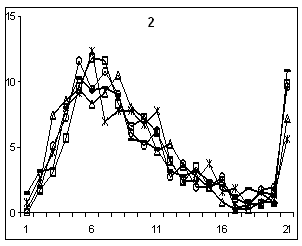
Рисунок 1. Изменения длин
экзонов (1, 2) в генах ядерного генома C. elegans. Экзоны в генах с одним (1) и пятью интронами (2). Ось
абсцисс – номер интервала длиной 20 н.; ось ординат – доля экзонов
соответствующей длины в каждом из интервалов в процентах. Обозначения: гены из хромосом: I и II (–●–); III (–□–); IV (–ж–) ; V (–○–); Х (–∆–).
Отметим, что гены с
одинаковым числом интронов, но из разных хромосом, отличались по количеству
интронов с длиной более 400 н. В разных группах генов хромосомы V доля таких интронов варьировала от
9,6 до 14,4%, тогда как в хромосомах I и II от 25,6 до 38,8%. В генах с разным
количеством интронов, независимо от хромосомной локализации генов, наблюдалось
больше всего интронов с длиной в интервале 40-60 н., с максимумом их числа при
длине 47-48 н. В интервале длин 40-60 н. доля интронов в генах хромосом I и II в среднем составляла 38%, в III - 41%, в IV - 38%, в V - 53% и в Х - 44%. То есть,
значительная доля интронов имела длину в этом интервале. Доля интронов с длиной в интервале 140-400 н. в
генах всех хромосом была близкой: 19,7±2,8 % - 26,9±2,3 %. Таким образом, перераспределение
длин интронов при увеличении числа экзонов в генах происходит путем уменьшения
доли интронов с длиной более 400 н. и увеличения их числа в интервале 40-60 н.,
без изменения доли интронов в промежуточных интервалах длин. Полученные
данные свидетельствуют, что гены с различным количеством интронов и
локализованные в разных хромосомах, отличаются друг от друга по своей
экзон-интронной организации. Тенденция увеличения числа интронов в гене, при
увеличении суммы длины экзонов, свидетельствует о корректирующей роли интронов
на пока неизвестные характеристики генов.
Длина экзонов и интронов
в генах не зависела от GC-содержания участка. Так, в участках 300 т.п.н. хромосом I и V, содержащих близкое число генов
(63÷65), но имеющих разное GC-содержание (39% и 34%), средняя
длина генов и соотношения длин экзонов и интронов в них не отличались. В генах,
локализованных в участках хромосом I и V с равным GC-содержанием (35%), но с различной их
плотностью, выявлено отличие по свойствам экзонов и интронов. Четыре участка с
высокой плотностью генов (УВПГ) содержали в среднем по 99 генов, а 4 УНПГ по
38. В хромосомах I и V средняя плотность генов составляла соответственно 61 и 73 гена на
участок длиной 300 т.п.н.
Средняя длина генов с
одним интроном равнялась в УВПГ и УНПГ 746 и 1106 н. при доле экзонов равной
соответственно 81 и 35%. В генах с двумя интронами средняя длина генов
составляла соответственно 1157 и 1896 н., а доли экзонов были 71 и 35%. В генах
с 6÷9 интронами длина генов равнялась 2855 и 7696 н., а экзоны
составляли 53 и 23% от длины генов. Для генов с 10 и более интронами отмеченная
тенденция сохранялась. Несмотря на значительное отличие в плотности генов в
участках, доля ДНК занимаемой белок-кодирующими генами в УВПГ и УНПГ была
близкой и составляла в среднем соответственно 47 и 49%. Средняя длина интронов
в генах УВПГ была меньше таковой в генах УНПГ в 4,3 раза. Несмотря на
значительные отличия длины интронов в генах C. еlegans, локализованных в участках ДНК с различной плотностью генов,
длина суммы экзонов была близкой в группах генов с одинаковым числом интронов.
Таким образом, экзон-интронная организация генов C. еlegans зависит от плотности генов в
участках ДНК ядерного генома.
Одно из свойств интронов
заключается в более низком GC-содержании по сравнению с GC-содержанием в экзонах, что приводит
к уменьшению его в генах (таблица 1). Другой характеристикой генов является
разность отношений частот использования нуклеотидов: fA/fT–fC/fG. Этот показатель отражает отклонение
fA, fT, fC, fG от отношений fA=fT и fC=fG, характерных согласно второму правилу Чаргаффа для протяженной
однонитевой ДНК. В экзонах величина fA/fT–fC/fG является положительной для генов,
кодирующих гидрофильные белки (fR-0,5>0), и отрицательной для генов, кодирующих гидрофобные
белки (fR-0,5<0), причем она существенно отличается от нуля. На наш взгляд
интроны, приближая величину fA/fT–fC/fG к нулю, оптимизируют нуклеотидный
состав гена. В генах без интронов величина fA/fT–fC/fG для генов гидрофильных и гидрофобных
белков равнялась соответственно 0,342 и -0,357. В генах с разным числом
интронов происходило снижение абсолютной величины fA/fT–fC/fG под влиянием интронов. Наиболее ярко
это видно для генов кодирующих гидрофильные белки, у которых величина fA/fT–fC/fG положительна. Для генов гидрофобных
белков заметное уменьшение величины fA/fT–fC/fG проявлялось в УВПГ.
Таблица 1. Изменения GC-содержания и величины fA/fT–fC/fG в экзонах и генах в УВПГ и УНПГ из
хромосом I и V C. еlegans
|
Число интронов |
fR |
Длина экзоновн. |
fS |
fR-0,5 |
fA/fT–fC/fG |
Длина гена, н. |
fS |
fR-0,5 |
fA/fT–fC/fG |
|
Гены без интронов |
>0,5 <0,5 |
780 652 |
0,437 0,470 |
0,041 -0,037 |
0,342 -0,357 |
|
|
|
|
|
Гены из УНПГ |
|||||||||
|
1-2 |
>0,5 <0,5 |
608 469 |
0,456 0,443 |
0,040 -0,038 |
0,322 -0,312 |
1911 1159 |
0,396 0,387 |
0,015 -0,019 |
0,110 -0,130 |
|
3-5 |
>0,5 <0,5 |
1015 903 |
0,451 0,420 |
0,036 -0,025 |
0,302 -0,192 |
3352 2171 |
0,377 0,389 |
0,014 -0,020 |
0,110 -0,191 |
|
6-9 |
>0,5 <0,5 |
1750 2148 |
0,451 0,478 |
0,031 -0,006 |
0,296 -0,054 |
6953 11162 |
0,368 0,342 |
0,007 -0,004 |
0,046 -0,069 |
|
≥10 |
>0,5 <0,5 |
4835 2535 |
0,454 0,470 |
0,030 -0,008 |
0,248 -0,047 |
19948 24121 |
0,355 0,359 |
0,010 -0,012 |
0,083 -0,082 |
Интроны уменьшали и
относительное содержание пуриновых нуклеотидов (fR-0,5) в генах. В УНПГ интроны
значительно уменьшали величину fA/fT–fC/fG (таблица 1). В генах гидрофильных
белков из УВПГ интроны тоже существенно снижали величину fA/fT–fC/fG: с 0,398 до 0,205. Для всех генов
гидрофобных белков из УНПГ средняя длина равнялась 4532 н. и величина fA/fT–fC/fG была равна -0,146. Соответствующие
длины и величины fA/fT–fC/fG для экзонов этих генов равнялись 1100 н. и –0,212. Интроны в
генах гидрофобных белков из УВПГ изменяли величину fA/fT–fC/fG с –0,305 до –0,224. Средняя длина
генов и суммы экзонов составляла соответственно 1670 и 1043 н.
В протяженных участках
однонитевой ДНК (более 100 т.н.) различных эукариот величина fA/fT–fC/fG близка нулю. Такой баланс
соотношения частот встречаемости нуклеотидов (fA ≈ fT и fC ≈ fG) в протяженных участках достигается
за счет определенного сочетания частот встречаемости нуклеотидов в экзонах,
интронах и межгенных участках. Так, для хромосом I и V C. elegans, нами были проанализированы частоты
встречаемости нуклеотидов однонитевой ДНК разделенной последовательно на
участки по 3, 30 или 300 т.н. Показано, что средняя величина fA/fT–fC/fG для этих участков ДНК хромосомы I равна соответственно 0,0002, 0,003 и
0,004, а хромосомы V равна 0,0002, 0,004 и 0,004. Таким образом, интроны, составляя
существенную долю генов уменьшают величину fA/fT–fC/fG и тем самым оптимизируют
нуклеотидный состав ДНК.
Литература:
1. Bultrini E., Pizzi E.,
Giudice P., Frontani C. Pentamer vocabularies characterizing introns and
intro-like intergenic tracts from Caenorhabditis elegans and Drosophila
melanogaster // Gene. 2003. V. 304. P. 183-192.
2. Castillo-Davis C.I.,
Mekhedov S.L., Hart D.L., Koonin E.V., Kondrashov F.A. Selection for short
introns in highly expressed genes // Nature genetics. 2002. V.31. №4. P.
415-418.
3. Deutsch M., Long M. Intron-exon structures of eukaryotic model
organisms // Nucleic Acids Research. 1999. V. 27. №15. P. 3219-3228.
4. Castillo-Davis C.I.,
Mekhedov S.L., Hart D.L., Koonin E.V., Kondrashov F.A. Selection for short
introns in highly expressed genes // Nature genetics. 2002. V.31. №4. P.
415-418.